
«Ультимейты», «Экто» и обычные бензины — журнал За рулем
Сэкономить на заправке хочется каждому. Даже вполне обеспеченному человеку, разъезжающему, скажем, на БМВ или «Мерседесе» свежего модельного года. Но на задрипанную загородную АЗС, да еще с неизвестным брендом, такой человек не поедет ни за что: он выберет заведомо «правильную» АЗС с громким именем. Скажем, ВР или»Лукойл». И… задумается.
201110281734_no_copyright_bp_bak_2
Стоит ли переплачивать за топливо с приставками»Ультимейт» или «Экто»? Насколько обоснованны фирменные обещания увеличить мощность до 14,7%, поднять ускорение до 9,9% или снизить коррозию двигателя до нуля?
Для ответа на подобные вопросы мы приобрели на фирменных столичных АЗС образцы бензинов ВР и «Лукойл» с октановыми числами 95 и 98 — всего шесть образцов.
Материалы по теме
Материалы по теме
Купленные бензины по определению должны были оказаться должного качества — иначе всякие сравнения «Экто» и «не Экто» теряют смысл. Так и оказалось: проверка физико-химических параметров приобретенного топлива никаких неожиданностей не выявила. Все в порядке, действующие ГОСТы удовлетворены. Октановые числа — даже с превышением нормы, смол — мало.
Так и оказалось: проверка физико-химических параметров приобретенного топлива никаких неожиданностей не выявила. Все в порядке, действующие ГОСТы удовлетворены. Октановые числа — даже с превышением нормы, смол — мало.
Судя по заявлениям производителей топлив, бензины «Экто» и «Ультимейт» содержат моющие присадки. Насколько эффективно они работают и что от этого можно ждать?
Стандартный тест на моющую способность бензинов очень долог и трудоемок, но мы пойдем своим путем. Попробуем оценить саму тенденцию по изменению количества отложений при работе двигателя на бензинах с моющими присадками, а заодно проверить, как на это отреагирует мотор.
Эту процедуру проделали для каждого бензина — итог же оказался прост. Обычные топлива, которые мы брали в качестве образца для сравнения, слегка увеличили массу отложений. А вот топлива с моющими присадками дали обратный эффект — масса отложений стала уменьшаться. Причем результат и по «Ультимейтам», и по «Экто» был близким. Точность таких замеров невысока, поэтому 10…15% различия в результатах можно списать на погрешности, однако важна тенденция!
20140821_img_5456
Кстати, выявилось, что на 98-х бензинах моющие присадки работают чуть менее эффективно.
Материалы по теме
Материалы по теме
После «моющих» бензинов зафиксировали некоторый рост расхода воздуха. Значит, впускные каналы и клапаны стали пускать в двигатель больше смеси. Итог — рост мощности, снижение расхода топлива. Заметно снизилась токсичность по СН, но вот содержание оксидов азота немного увеличилось — сгорание пошло более эффективно, температуры поднялись — вот и результат. Впрочем, конфликт интересов между оксидами азота и остаточными углеводородами специалистам известен давно.
Тенденцию влияния современных «титулованных» топлив на мотор мы уяснили, однако пресловутых рекламных 14,7% снова не нашли. Но попробуем поставить еще один, уже итоговый, эксперимент, причем за пределами самой экспертизы — факультативно, так сказать…
Модельный эксперимент с искусственным загрязнением, который мы практиковали ранее, указал на тенденцию и дал возможность сравнения, но и только! А для выяснения реальных возможностей надо брать образец мотора с живой, натуральной грязью. Этого «добра» за время предыдущих испытаний накопилось много — им и воспользовались. Взяли 100 л одного из бензинов с моющей присадкой — «Лукойл — 95 Экто» и выработали его на этом моторе, сняв предварительно начальные характеристики. По ходу испытаний замерили динамику изменения параметров двигателя. Причем, по мере очистки, порог детонации сдвигался в сторону увеличения мощности — мы это тоже отслеживали.
Этого «добра» за время предыдущих испытаний накопилось много — им и воспользовались. Взяли 100 л одного из бензинов с моющей присадкой — «Лукойл — 95 Экто» и выработали его на этом моторе, сняв предварительно начальные характеристики. По ходу испытаний замерили динамику изменения параметров двигателя. Причем, по мере очистки, порог детонации сдвигался в сторону увеличения мощности — мы это тоже отслеживали.
Вот тут эффективность работы присадки проявилась более заметно. К концу цикла испытаний мощность подросла на 7,5%, а расход уменьшился в среднем на 8,4%. Токсичность упала только по остаточным углеводородам, оставаясь по остальным компонентам в пределах погрешности измерения. А если мотор будет еще более грязным? И бензина выработать побольше? Мы полагаем, что при этом рекламные цифры действительно могут стать досягаемыми.
Но вот что интересно! На начальном этапе работы нашего ну очень грязного мотора на хорошем бензине с моющей присадкой были замечены некоторые неприятности — и подергиваться стал, и токсичность заметно увеличилась. В чем дело? Мы полагаем, что загрязнения, смываемые со стенок топливной системы, попали в дозирующие элементы системы питания. Это и породило эффект. А когда двигатель слегка промылся, проявилась четкая тенденция к улучшению всех параметров. Вот так все непросто! И опять подтвердилась старая истина — не надо запускать болячки, то есть чистое всегда лучше грязного!
В чем дело? Мы полагаем, что загрязнения, смываемые со стенок топливной системы, попали в дозирующие элементы системы питания. Это и породило эффект. А когда двигатель слегка промылся, проявилась четкая тенденция к улучшению всех параметров. Вот так все непросто! И опять подтвердилась старая истина — не надо запускать болячки, то есть чистое всегда лучше грязного!
20141113_01
Так заливать моющее топливо в «грязный» мотор или нет? Мы рекомендуем переходить на хорошее постепенно. Сначала к обычному топливу добавить полбака бензина с моющей присадкой, затем, выработав бак на ¾, снова долить хорошего бензина. А потом уже заправляться только фирменным бензином. Заметим также, что на каком-то этапе придется поменять топливный фильтр, но это потребуется только в особо запущенных случаях.
Так что же в сухом остатке?
Во-первых, брендовые топлива действительно имеют определенные преимущества перед обычными. Это в основном проявляется из-за улучшенных моющих свойств подобных бензинов. Однако увидеть это удается не сразу, а при постоянном их применении. Но если бензин позиционируется как соответствующий Евро-3 и выше, то содержать моющие присадки он просто обязан.
Однако увидеть это удается не сразу, а при постоянном их применении. Но если бензин позиционируется как соответствующий Евро-3 и выше, то содержать моющие присадки он просто обязан.
Во-вторых, основные преимущества таких бензинов проявляются все-таки на современных впрысковых моторах, но они для них ведь и создавались! Дело в том, что струя топлива из форсунки в таком моторе позволяет лучше использовать моющие свойства присадок, чем в карбюраторном. Поэтому ждать эффекта для «старичка» придется значительно дольше.
В-третьих, заявленных 14,7% прироста мощности мы не нашли. Но если использовать все возможности, которые предоставляют бензины с более высокими октановыми числами и хорошей моющей способностью, а в качестве подопытного кролика взять что-то сильно грязное, то такие цифры могут быть вполне реальными. Но речь, скорее, не об увеличении мощности, а об ее восстановлении.
И, наконец, в-четвертых: уж не настолько титулованные бензины дороже обычных… Да еще при постоянно растущей стоимости самых простых топлив?
Подробнее — по ссылке.
[Все об автомобильном бензине] [Бензин: Евро, классы и EN 228] [Экспертизы и исследования бензинов]
[
Сэкономить на заправке хочется каждому. Даже вполне обеспеченному человеку, разъезжающему, скажем, на БМВ или «Мерседесе» свежего модельного года. Но на задрипанную загородную АЗС, да еще с неизвестным брендом, такой человек не поедет ни за что: он выберет заведомо «правильную» АЗС с громким именем. Скажем, ВР или »Лукойл». И… задумается.
«Ультимейты», «Экто» и обычные бензиныЛУКОЙЛ — Инновационное топливо ЭКТО
Больше плюсов в каждом километре с Экто PLUS от ЛУКОЙЛ
Экто PLUS — это улучшенное топливо от ЛУКОЙЛ, с которым достигается
максимальная мощность двигателя, снижается расход топлива и обеспечивается
защита двигателя от износа.
За счет чего это происходит?
Преимущества обновленного топлива Экто Plus обеспечиваются высоким качеством базового топлива стандарта Евро-5 компании ЛУКОЙЛ, а также благодаря новому пакету присадок Keropur®️ ECTO, обеспечивающему высокий эффект по очистке и поддержанию чистоты топливной системы. Пакет присадок Keropur®️ ECTO покрывает детали системы впрыска двигателя защитной плёнкой, предотвращает образование отложений во впускной системе, поддерживает в чистоте впускные клапаны и топливные инжекторы.
Для чего нужен пакет присадок Keropur®️ ECTO
Стандартные топлива, соответствующие государственным стандартам, могут образовывать отложения. Отложения на впускных клапанах, в топливных форсунках и в камере сгорания могут оказать серьезное негативное влияние на работоспособность двигателя:
- нарушить оптимальное соотношения топливо/воздух, что приведет к неполному сгоранию смеси;
увеличить расход топлива;
увеличить токсичность отработавших газов;
снизить мощность;
негативно повлиять на управляемость;
увеличить шумы и вибрации.


Преимущества нового пакета Keropur® ECTO
- Keropur® ЕСТО представляет собой полностью совместимый сбалансированный пакет;
- Моющая способность является ключевым преимуществом, который улучшает качество топлива;
- Кроме моющего компонента он содержит модификатор трения, деэмульгатор и высокоэффективный ингибитор коррозии, который защищает от коррозионного разрушения системы подачи топлива, резервуары для хранения, топливопроводы и оборудование заправочных станций;
- Полностью синтетическая формула Keropur® ЕСТО позволяет контролировать образование отложений в камере сгорания:
· Покрывает детали системы впрыска двигателя защитной плёнкой;
· Предотвращает образование отложений и поддерживает в чистоте впускные клапаны и топливные инжекторы;
· Удаляет существующие отложения.

Основные преимущества обновленного топлива Экто PLUS
+11% экономия топлива
Модификатор трения в пакете Keropur®️ ECTO накапливается в моторном масле и приводит к снижению силы трения между всеми деталями, находящимися с ним в контакте, этот эффект является кумулятивным и длительным по времени. Также расход топлива снижается благодаря эффективному горению.
Среднее значение по сравнению с бензином АИ-95-К5 производства ООО «ЛУКОЙЛ-Нижегороднефтеоргсинтез» без присадки.
+50% защита двигателя от износа
Пакет присадок Keropur®️ ECTO покрывает детали системы впрыска двигателя защитной плёнкой, предотвращает образование отложений и поддерживает в чистоте впускные клапаны и топливные инжекторы.
По сравнению с бензином АИ-95-К5 производства ООО «ЛУКОЙЛ-Нижегороднефтеоргсинтез» без присадки.
+13.8% увеличение мощности
Модификатор трения в пакете Keropur® ECTO обеспечивает снижение трения, и прирост КПД двигателя достигается за счет создания тонкой пленки на стенках камеры сгорания (этот эффект наблюдается сразу после начала работы мгновенный эффект).
Происходит оптимизация процесса образования топливно-воздушной смеси и обеспечение ее максимально эффективного сгорания.
До +13,8 % по сравнению с бензином АИ-95-К5 производства ООО «ЛУКОЙЛ-Нижегороднефтеоргсинтез» без присадки.
Преимущества топлива доказаны во время испытаний
Всего выполнено более 80 тестов в центре моторных испытаний «Competence Center
Fuels and Lubes» (CCFL) в Германии. Испытания на динамометрических стендах
проведены в независимом испытательном центре «COMPLIANCE AND RESERCH SERVICES,
INC.», в Нью-Джерси, США.
Вас может также заинтересовать
95 Экто: линейка бензинов от Лукойл
Свойства и особенности топлива Отличия Экто от Евро
Российские производители бензина теперь следят не только за соблюдением его октанового числа, но и его соответствием общепринятым европейским стандартам. В базовый состав топлива вводятся оригинальные добавки.
В базовый состав топлива вводятся оригинальные добавки.
Свои оригинальные марки бензина предлагают компании Газпромнефть (G-Drive), Роснефть (Пульсар) и др.
Фирменным топливом Лукойл является бензин Экто. Рассмотрим его особенности, достоинства и отличия от Евро.
Особенности топлива Экто
Основная особенность автомобильных бензинов Экто от компании Лукойл – в составе присадок, эффективность которых была предварительно протестирована на оборудовании авторитетной британской фирмы «Tickford Power Train Test Ltd».
В ходе испытаний оценивались:
- Уровень вредных выхлопов
- Детонационные характеристики
- Текущая мощность двигателя
- Удельный расход топлива
Имеются данные, что используемые в топливе Экто добавки позволяют поднять уровень эксплуатационных свойств данного бензина до уровня Евро-5. Это позволяет автомобилистам, которые пользуются услугами фирменных АЗС от Лукойл, без проблем посещать страны Евросоюза.
Это позволяет автомобилистам, которые пользуются услугами фирменных АЗС от Лукойл, без проблем посещать страны Евросоюза.
Виды топлива Экто
Линейка рассматриваемого топлива включает в себя 3 разновидности бензина:
- Экто-92
- Экто-95
- Экто-100 (Экто Plus)
Фактическое октановое число бензина марки Экто-92 составляет не менее 95, а Экто-95 – 97 единиц.
Все виды топлива Экто гарантируют отсутствие коррозионной опасности для стальных деталей, очищение инжектора и увеличение ресурса двигателя. Экто Плюс, по заявлению Лукойл, снижает расход топлива на 5… 6 %.
Производитель отмечает также, что предлагаемая линейка ориентирована, прежде всего, на автомобили европейских производителей – Porsche, BMW и некоторые другие.
Рассмотрим свойства самого популярного в серии Экто бензина с октановым числом 95.
Свойства Экто 95
Бензин Экто-95 соответствует стандарту Евро-3. Перечислим основные свойства этого топлива:
- Низкое содержание ароматических углеводородов
- Доля кислорода – до 2,7 %
- Содержание оксигенатов – до 15 %
- Цетановый индекс – от 46
- Цетановое число – 51
- Точка воспламенения – 55 °С
Экто-95 отличается пониженный содержанием бензола, что гарантирует его низкую токсичность и уменьшает интенсивность нагарообразования в двигателе.
Преимущества Экто 95
Главными преимуществами бензина 95-Экто, которые выделяют его на фоне других, заключаются в:
- Повышении надежности силового агрегата, а также продлении срока его службы
- Обеспечении максимальной мощности автомобиля
- Наличии специализированных присадок, защищает ключевые узлы от коррозии
- Снижение уровня детонации, приводящему к более тихой и бесшумной работе двигателя
- Повышении срока службы системы впрыска
- Минимизации осадка
- Уменьшении расхода топлива
Этими же достоинствами обладает дизельное топливо Экто от Лукойл. Как утверждают разработчики, его использование позволяет уменьшить расход топлива фактически на 3,5 %. Наличие моющих присадок существенно уменьшает количество налета, что делает работу силового агрегата значительно тише.
Как утверждают разработчики, его использование позволяет уменьшить расход топлива фактически на 3,5 %. Наличие моющих присадок существенно уменьшает количество налета, что делает работу силового агрегата значительно тише.
Чем Экто отличется от Евро?
Психология многих водителей понятна: имея хороший автомобиль, они хотят ездить на фирменном бензине, пусть даже с переплатой. Чтобы оценить реальные преимущества бензинов Лукойл Экто от Евро, были проведены сравнительные испытания. Они показали следующее.
Количество смолистых компонентов в топливе Экто действительно снижено (относительно параметров, установленных для бензинов класса Евро-4).
Наличие моющих присадок на самом деле увеличивает мощность двигателя (особенно это актуально для топлива с повышенным октановым числом). В результате снижается процент выделяемых углеводородов. При этом процент окислов азота в выхлопных газах увеличивается, что связано с ростом температуры в камере сгорания.
В топливе Евро нет моющих присадок. Именно поэтому его, а не Экто, рекомендуется использовать в старых автомобилях с большим пробегом. Это объясняется тем, что наличие очищающих добавок в бензине Экто может привести к засорению топливного фильтра отслоившимися загрязнениями.
Показатели Экто в плане экологичности улучшены по сравнению с Евро: содержание серы и бензола понижено в несколько раз, поэтому можно сказать, что Экто является улучшенной версией Евро-3/
BP Ultimate и ЛУКОЙЛ “ЭКТО” – лить или не лить? :: Autonews
В последнее время владельцам автомобилей тяжело: чем заправлять своих верных железных друзей?! Отовсюду несётся реклама самых новых, самых лучших, самых полезных для двигателя сортов топлива. В такой ситуации сделать выбор совсем не просто, особенно не обладая достаточным набором знаний. Попробуем разобраться, что к чему.
Попробуем разобраться, что к чему.Надо отдать должное компаниям ТНК-BP и ЛУКОЙЛ, реклама их продукции очень впечатляет. Ну кто, скажите, в здравом уме откажется от практически полной очистки двигателя, повышения экономичности, да еще и увеличения мощности на 10-15 процентов?! Но далеко не факт, что всё так уж безоблачно и приятно.
По официальной информации BP Ultimate 98 демонстрирует увеличение мощности до 14,7 % и улучшение ускорения до 9,9 %, по сравнению с двигателями потребляющими обычный Аи-95. Много говорится и об очищающих свойствах, о том, что со временем удаляются даже уже имеющиеся отложения. Так же производители утверждают, что Ultimate значительно меньше загрязняет окружающую среду. Наверняка, многие видели впечатляющие фотографии впускных клапанов, одни, снятые с двигателя, регулярно работавшего на обычном топливе, другие, с мотора, потреблявшего Ultimate. Естественно, первые выглядят ужасно, а вторые сверкают чистотой и новизной. Ну и конечно, стоит оценить фразу о снижении потребления топлива. В среднем можно сэкономить до 20 км с бака.
В среднем можно сэкономить до 20 км с бака.
Рекламная компания “ЭКТО” выглядит скромнее. Тут обещают “всего лишь “ 100% от заявленной мощности мотора, его более надёжную работу, увеличение интервала замены масла, снижение шума и вибрации, и т.д. Плюс “ЭКТО” содержит в пять раз меньше бензола и в 3 раза меньше серы и, соответственно, является более экологичным. Причем, тут же имеется честное предупреждение о том, что если двигатель имеет большой пробег, то применение такого топлива может привести к некоторым отклонениям в его работе, что является обычной практикой при использовании такого бензина.
Как бы в пику всем дифирамбам, которые поются этим двум суперсортам топлива, то тут, то там натыкаешься на негативные отзывы об Ultimate и “ЭКТО”. Кто-то уже после пары заправок отправился на переборку двигателя, кому-то пришлось срочно менять свечи, у кого-то машина просто отказалась заводиться. Кому-то по секрету сказали механики, что этот бензин хуже, чем Аи-92 или даже Аи-80. Скорее всего, всё это полный бред. Механики – оказались не самыми сведущими в вопросах топлива, а поломки – просто совпадения. Если только топливо не было залито в какой-нибудь глухой деревне, а слова Ultimate или “ЭКТО” не были старательно выведены маркером на заправочном автомате. Хотя очень многих такие отзывы убеждают в том, что нельзя заливать в свои автомобили ни Ultimate, ни “ЭКТО”.
Скорее всего, всё это полный бред. Механики – оказались не самыми сведущими в вопросах топлива, а поломки – просто совпадения. Если только топливо не было залито в какой-нибудь глухой деревне, а слова Ultimate или “ЭКТО” не были старательно выведены маркером на заправочном автомате. Хотя очень многих такие отзывы убеждают в том, что нельзя заливать в свои автомобили ни Ultimate, ни “ЭКТО”.
Что касается разительного увеличения мощности, то после экспериментов, проведённых известными автоэкспертами, выяснилось, что на отечественные машины это не распространяется. Что касается иномарок, то и тут всё не так уж просто. Если прирост мощности и есть, то явно не 14%. А вот с экономичностью дело обстоит получше. Расход, абсолютно точно не увеличивается, даже уменьшается, но достаточно незначительно.
Часто водители полагают, что тот же Ultimate и “ЭКТО” – это всего лишь качественно топливо, соответствующее своему октановому числу. При этом на одной и той же заправке продаются старое – некачественное топливо, и новое – качественное. Люди, которые заботятся о своем автомобиле, не жалеют денег, чтобы избежать проблем. Поэтому наличие такого бензина можно считать поднятием репутации АЗС. Переплачивая рубль-другой за литр топлива, человек надеется, что в нём не будет вредных присадок, которые способны в момент уничтожить свечи.
Люди, которые заботятся о своем автомобиле, не жалеют денег, чтобы избежать проблем. Поэтому наличие такого бензина можно считать поднятием репутации АЗС. Переплачивая рубль-другой за литр топлива, человек надеется, что в нём не будет вредных присадок, которые способны в момент уничтожить свечи.
Однако, не стоит сразу относиться к этим видам топлива, как к обману. В них содержатся моющие компоненты, которые действительно позволяют избежать некоторых неприятностей. Пожалуй, в большей степени это относится к автомобилям иностранного производства. Использование моющих присадок позволит избежать перебоев в работе двигателя, забивки форсунок. Что, соответственно, поможет сэкономить некоторое количество денег и большое количество времени.
Николай Загвоздкин
В Беларуси расширена реализация нового брендированного топлива АИ-95 ЭКТО Plus
«ЛУКОЙЛ» представляет инновационное топливо «АИ-95-К5 ЭКТО Plus» теперь и в Республике Беларусь!
«ЭКТО» — это линейка инновационных топлив, соответствующих европейскому уровню качества. Топливо «АИ-95-К5 ЭКТО Plus» создано с учётом самых современных технологий и с добавлением многофункционального пакета топливных присадок последнего поколения. Актуально, что при его использовании снижается выброс углекислого газа в окружающую среду.
Топливо «АИ-95-К5 ЭКТО Plus» создано с учётом самых современных технологий и с добавлением многофункционального пакета топливных присадок последнего поколения. Актуально, что при его использовании снижается выброс углекислого газа в окружающую среду.
«АИ-95-К5 ЭКТО Plus» — чтобы автолюбители и профессионалы получали ещё больше удовольствия от вождения.
Детально отметим достоинства «АИ-95-К5 ЭКТО Plus»:
- — Увеличение мощности двигателя при использовании топлива «ЭКТО Plus» подтверждено испытаниями независимого британского центра Tickford Power Train Test Ltd. Специальный модификатор трения снижает механическое трение в цилиндро-поршневой группе, обеспечивая великолепную динамику автомобиля и повышая КПД двигателя.
- — Высочайшее качество «ЭКТО Plus» соответствует международному экологическому стандарту Евро-5, уменьшает нагрузку на систему нейтрализации отработанных газов, улучшает работу автомобиля и заботится об окружающей среде за счёт низкого содержания веществ в выхлопе.
- — «ЭКТО Plus» повышает эффективность работы двигателя, одновременно очищая его. Моющий компонент предотвращает образование отложений на деталях двигателя, поддерживает в чистоте инжектор и впускные клапаны, обеспечивая работу двигателя в соответствии с оптимальными заводскими настройками. Специальные компоненты дают дополнительный эксплуатационный эффект: защищают двигатель от образования нагара и от коррозии топливной системы.
- — Применение «ЭКТО Plus» снижает расход топлива и даёт дополнительный пробег с каждого полного бака бензина. Это значит, что при выборе в пользу «АИ-95-К5 ЭКТО Plus» необходимо реже заезжать на заправку.

«ЭКТО Plus» способно наполнить двигатель автомобиля неудержимой энергией, заставляя его пульс биться в такт вашим желаниям.
В 2020 году на территории Республики Беларусь началась продажа бензина «АИ-95 ЭКТО PLUS» в г. Минске и Минской области, г. Бресте и Брестской области. С ноября 2021 года сеть АЗС «ЛУКОЙЛ» расширяет географию по продаже брендового бензина «АИ-95 ЭКТО PLUS» — он реализуется во всех областных городах Беларуси на 57 АЗС.
Процесс производства бензина «АИ-95 ЭКТО PLUS» осуществляется прямо на нефтебазах ИООО «ЛУКОЙЛ Белоруссия». Процесс смешения контролируется компьютером, многофункциональная присадка, добавляемая в базовое топливо, привозится в специальных бочках и в ходе производства добавляется уже при заправке автоцистерны через специальную форсунку, дозирующую заданную пропорцию состава. У компании ИООО «ЛУКОЙЛ Белоруссия» имеются собственные независимые лаборатории в Беларуси, которые позволяют отслеживать качество выпускаемого топлива. Это важнейшая составляющая для компании, ведь стабильное качество — залог успешного бизнеса.
Вы можете испытать все свойства нового продукта по цене базового топлива, а также с ноября 2021 года на АЗС «ЛУКОЙЛ» в Беларуси можете поучаствовать в акции «ЭКТО ПЛЮС ВЫГОДА» и выиграть подарочный сертификат.
Пробуйте, заправляйтесь, получайте призы! Подробности на сайте.
Заправляя свой автомобиль топливом «АИ-95-К5 ЭКТО Plus», вы наслаждаетесь потрясающей динамикой вождения.
«ЭКТО Plus» заводит вашу машину!
Пермский УФАС проверит «дочку» «Лукойла» после резкого повышения цен на бензин | Рынок нефтепродуктов
Дизельное топливо подорожало на 1,6 рубля за литр, бензин марок АИ-92 и ЭКТО-92, АИ-95 — на 64 копейки
Управление ФАС по Пермскому краю начало проверку ООО «Лукойл-Уралнефтепродукт», занимающего в регионе доминирующее положение на розничном рынке нефтепродуктов. В ведомстве сомневаются в обоснованности резкого повышения цен на АЗС компании, произошедшего вопреки снижению биржевых котировок на топливо, сообщает в понедельник пресс-служба УФАС.
«В целях установления причин ноябрьского повышения цен антимонопольным ведомством в адрес ООО «Лукойл-Уралнефтепродукт» оперативно подготовлен и направлен запрос необходимых пояснений и документов, которые позволят объективно оценить динамику затрат на закуп нефтепродуктов и динамику розничных цен в других регионах присутствия. В случае выявления в действиях ООО «Лукойл-Уралнефтепродукт» признаков установления монопольно высокой цены будет возбуждено дело о нарушении антимонопольного законодательства и приняты все необходимые меры реагирования», — передают в сообщении слова заместитель руководителя пермского УФАС Дмитрия Шилова.
В случае выявления в действиях ООО «Лукойл-Уралнефтепродукт» признаков установления монопольно высокой цены будет возбуждено дело о нарушении антимонопольного законодательства и приняты все необходимые меры реагирования», — передают в сообщении слова заместитель руководителя пермского УФАС Дмитрия Шилова.
Отмечается, что на АЗС ООО «Лукойл-Уралнефтепродукт» в Пермском крае на выходных во второй раз за неделю резко выросли цены на бензин. Суммарный рост по маркам АИ-92 и ЭКТО-92, АИ-95 составил 64 копейки за литр, по дизельному топливу — 1,60 рубля за литр. За год рост общий уровень розничных цен вырос на 7-8%.
При этом рост произошел на фоне снижения территориальных биржевых индексов нефтепродуктов. Ведомство приводит в пример данные Санкт-Петербургской международной товарно-сырьевой биржи, где за последние два месяца стоимость бензинов АИ-92 (индикатор REG) снизилась на 13,25%, бензинов АИ-95 (индикатор PRM) снизилась на 15,42%.
«Лукойл-Уралнефтепродукт» — один из крупнейших межрегиональных сбытовых операторов ПАО «Лукойл». Предприятие создали в 2004 году, является 100% дочерней структурой компании. «Лукойл-Уралнефтепродукт» занимается реализацией топлива, масел, сопутствующих товаров, хранением нефтепродуктов и эксплуатацией нефтебаз в республиках Башкортостан и Татарстан, на территории Оренбургской, Самарской, Тюменской, Челябинской, Курганской, Свердловской, Кировской и Ульяновской областей, Удмуртской республики, ХМАО, ЯНАО и Пермском крае. В состав предприятия входят более 600 автозаправочных комплексов и 12 нефтебаз.
Предприятие создали в 2004 году, является 100% дочерней структурой компании. «Лукойл-Уралнефтепродукт» занимается реализацией топлива, масел, сопутствующих товаров, хранением нефтепродуктов и эксплуатацией нефтебаз в республиках Башкортостан и Татарстан, на территории Оренбургской, Самарской, Тюменской, Челябинской, Курганской, Свердловской, Кировской и Ульяновской областей, Удмуртской республики, ХМАО, ЯНАО и Пермском крае. В состав предприятия входят более 600 автозаправочных комплексов и 12 нефтебаз.
Пермский УФАС проверит «дочку» «Лукойла» после резкого повышения цен на бензин — Экономика и бизнес
ПЕРМЬ, 29 ноября. /ТАСС/. Управление ФАС по Пермскому краю начало проверку ООО «Лукойл-Уралнефтепродукт», занимающего в регионе доминирующее положение на розничном рынке нефтепродуктов. В ведомстве сомневаются в обоснованности резкого повышения цен на АЗС компании, произошедшего вопреки снижению биржевых котировок на топливо, сообщает в понедельник пресс-служба УФАС.
«В целях установления причин ноябрьского повышения цен антимонопольным ведомством в адрес ООО «Лукойл-Уралнефтепродукт» оперативно подготовлен и направлен запрос необходимых пояснений и документов, которые позволят объективно оценить динамику затрат на закуп нефтепродуктов и динамику розничных цен в других регионах присутствия. В случае выявления в действиях ООО «Лукойл-Уралнефтепродукт» признаков установления монопольно высокой цены будет возбуждено дело о нарушении антимонопольного законодательства и приняты все необходимые меры реагирования», — передают в сообщении слова заместитель руководителя пермского УФАС Дмитрия Шилова.
Отмечается, что на АЗС ООО «Лукойл-Уралнефтепродукт» в Пермском крае на выходных во второй раз за неделю резко выросли цены на бензин. Суммарный рост по маркам АИ-92 и ЭКТО-92, АИ-95 составил 64 копейки за литр, по дизельному топливу — 1,60 рубля за литр. За год рост общий уровень розничных цен вырос на 7-8%.
При этом рост произошел на фоне снижения территориальных биржевых индексов нефтепродуктов. Ведомство приводит в пример данные Санкт-Петербургской международной товарно-сырьевой биржи, где за последние два месяца стоимость бензинов АИ-92 (индикатор REG) снизилась на 13,25%, бензинов АИ-95 (индикатор PRM) снизилась на 15,42%.
«Лукойл-Уралнефтепродукт» — один из крупнейших межрегиональных сбытовых операторов ПАО «Лукойл». Предприятие создали в 2004 году, является 100% дочерней структурой компании. «Лукойл-Уралнефтепродукт» занимается реализацией топлива, масел, сопутствующих товаров, хранением нефтепродуктов и эксплуатацией нефтебаз в республиках Башкортостан и Татарстан, на территории Оренбургской, Самарской, Тюменской, Челябинской, Курганской, Свердловской, Кировской и Ульяновской областей, Удмуртской республики, ХМАО, ЯНАО и Пермском крае. В состав предприятия входят более 600 автозаправочных комплексов и 12 нефтебаз.
Экто-АДФ-рибозилтрансферазы (АРТ): новые участники клеточной коммуникации и передачи сигналов
Название: Экто-АДФ-рибозилтрансферазы (АРТ): новые участники клеточной коммуникации и передачи сигналов
Объем: 11 Выпуск: 7
Автор(ы): Мишель Семан, Сахил Адриуш, Фридрих Хааг и Фридрих Кох-Нольте
Принадлежность:
- Laboratoired’Immunodifferenciation, EA1556, Tour 54, CP 7124, 2 место Jussieu75251 Paris Cedex 05, Франция. ,Франция
Ключевые слова: экто-адп-рибозилтрансферазы, поли(адп-рибозил)трансферазы, арт-токсины, клеточная коммуникация
Abstract: Экто-АДФ-рибозилтрансферазы млекопитающих (ARTs) представляют собой семейство структурно родственных белков, экспрессируемых на клеточной поверхности или секретируемых во внеклеточном компартменте. Используя НАД+ в качестве субстрата, они переносят АДФ-рибозные группы на белки-мишени.В отличие от внутриклеточных поли(АДФ-рибозил)трансфераз (PARP), эти ферменты переносят одну ADPR и, таким образом, являются моно-АРТ. Пять паралогов (ART1-5) были клонированы, но только четыре из них экспрессируются у человека из-за дефектного гена ART2 и шесть у мыши в результате дупликации гена ART2. Недавнее определение кристаллической структуры крысиного ART2 выявило гомологию с бактериальными токсинами ART и обеспечило молекулярную основу для понимания специфичности ART2 в отношении их мишеней. Комбинация различных технологических подходов показывает, что экто-АРТ экспрессируются в различных тканях с привилегированными участками, такими как сердце и скелетные мышцы для АРТ1, Т-лимфоциты для АРТ2 или яички для АРТ5. Это также указывает на то, что экспрессия ART строго регулируется. АДФ-рибозилирование белков-мишеней на клеточных поверхностях или циркулирующих в жидкостях организма приводит к обратимым посттрансляционным модификациям, которые могут ингибировать мишени, как известно для бактериальных АРТ, или активировать их, как в перекрестных помехах между мышиным ART2 и цитолитическим рецептором P2X7 на Т-лимфоциты.Активность АРТ во внеклеточном компартменте обеспечивает сложные регуляторные механизмы клеточной коммуникации. Это указывает на то, что экто-АРТ являются новыми кандидатами для таргетной терапии.
Комбинация различных технологических подходов показывает, что экто-АРТ экспрессируются в различных тканях с привилегированными участками, такими как сердце и скелетные мышцы для АРТ1, Т-лимфоциты для АРТ2 или яички для АРТ5. Это также указывает на то, что экспрессия ART строго регулируется. АДФ-рибозилирование белков-мишеней на клеточных поверхностях или циркулирующих в жидкостях организма приводит к обратимым посттрансляционным модификациям, которые могут ингибировать мишени, как известно для бактериальных АРТ, или активировать их, как в перекрестных помехах между мышиным ART2 и цитолитическим рецептором P2X7 на Т-лимфоциты.Активность АРТ во внеклеточном компартменте обеспечивает сложные регуляторные механизмы клеточной коммуникации. Это указывает на то, что экто-АРТ являются новыми кандидатами для таргетной терапии.
Зависимость от времени и температуры связывания 19-kDa GP Ecto с липосомами….
Контекст 1
. .. ген филовируса, для которого существует вирус Эбола (EBOV), в настоящее время не является чрезвычайно одобренной смертельной вакциной — или противовирусные средства (18).Проникновение вируса в клетки опосредуется GP, единственным видом гликопротеина, присутствующим на поверхности вириона (32). GP представляет собой слитый белок класса I (22, 53), который обеспечивает связывание с клеточной поверхностью и слияние с внутриклеточной мембраной. Подобно многим другим слитым белкам класса I (например, гемагглютинину вируса гриппа), нативный GP представляет собой тример гетеродимеров, состоящий из рецептор-связывающей субъединицы (GP1) и слитой субъединицы (GP2) (31). Две субъединицы возникают в результате расщепления полипептида-предшественника, когда GP проходит компартмент Гольджи, и остаются связанными посредством дисульфидной связи и нековалентных взаимодействий.GP1 представляет собой большую субъединицу (кажущаяся молекулярная масса 130 кДа), которая включает гликановый кэп, содержащий 4 N-связанных сайта присоединения углеводов и сильно O-гликозилированный муциноподобный домен. Муциноподобный домен необязателен для инфицирования клеток в культуре; псевдовирионы и вирусоподобные частицы, несущие генно-инженерный GP с делецией муциноподобного домена (GP ⌬), полностью способны к проникновению и инфицированию (17, 29, 46). Два богатых углеводами домена GP1 расположены поверх головного домена, который содержит рецептор-связывающую область (RBR) (5, 17, 29, 35, 39).В свою очередь, головной домен GP1 расположен на основании, которое экстенсивно взаимодействует с субъединицей слияния (GP2), в которой находится петля слияния (26). Эндосомальные протеазы катепсин B (Cat B) и катепсин L (Cat L) необходимы для проникновения EBOV в цитоплазму и, следовательно, для инфицирования клеток-хозяев (10, 29, 45). Cat B и Cat L расщепляют GP внутри неструктурированной  13-  14 петли GP1, генерируя промежуточное соединение 20-kDa, а затем ключевое ядро 19-kDa (17, 25, 45). Процессинг, по-видимому, проходит через промежуточное соединение ϳ 50 кДа, которое напоминает GP ⌬.Во время этих массовых событий расщепления GP2 полностью защищен и остается дисульфидно связанным с GP1. Процессинг до GP 19 кДа удаляет муциноподобный домен, гликановый кэп и внешнюю β-цепь ( 14) головного домена (GP1). Район связывания рецептора (RBR) и основание GP1, а также весь GP2 остаются интактными. Мы определили расщепление GP1 катепсином как стадию праймирования, поскольку мы предположили, что GP2 остается зажатым в ядре 19 кДа (45). Доказательства в поддержку этого предложения включают наблюдение, что проникновение псевдовирионов, несущих 19-кДа GP, блокируется как бафиломицином, так и E64(d) (45, 54), предполагая, что низкий pH и чувствительный к E64 процесс необходимы для индукции 19- kDa GP для обеспечения слияния и проникновения.Первая не исключающая друг друга гипотеза (45), объясняющая эти находки, заключается в том, что праймирование ядра 19-kDa усиливает реакцию GP на триггер(ы) слияния. Во-вторых, расщепление до 19 кДа усиливает способность GP связываться с эндосомальным рецептором (29). Большинство вирусных слитых белков находятся на соответствующих поверхностях вирионов в метастабильном состоянии.
.. ген филовируса, для которого существует вирус Эбола (EBOV), в настоящее время не является чрезвычайно одобренной смертельной вакциной — или противовирусные средства (18).Проникновение вируса в клетки опосредуется GP, единственным видом гликопротеина, присутствующим на поверхности вириона (32). GP представляет собой слитый белок класса I (22, 53), который обеспечивает связывание с клеточной поверхностью и слияние с внутриклеточной мембраной. Подобно многим другим слитым белкам класса I (например, гемагглютинину вируса гриппа), нативный GP представляет собой тример гетеродимеров, состоящий из рецептор-связывающей субъединицы (GP1) и слитой субъединицы (GP2) (31). Две субъединицы возникают в результате расщепления полипептида-предшественника, когда GP проходит компартмент Гольджи, и остаются связанными посредством дисульфидной связи и нековалентных взаимодействий.GP1 представляет собой большую субъединицу (кажущаяся молекулярная масса 130 кДа), которая включает гликановый кэп, содержащий 4 N-связанных сайта присоединения углеводов и сильно O-гликозилированный муциноподобный домен. Муциноподобный домен необязателен для инфицирования клеток в культуре; псевдовирионы и вирусоподобные частицы, несущие генно-инженерный GP с делецией муциноподобного домена (GP ⌬), полностью способны к проникновению и инфицированию (17, 29, 46). Два богатых углеводами домена GP1 расположены поверх головного домена, который содержит рецептор-связывающую область (RBR) (5, 17, 29, 35, 39).В свою очередь, головной домен GP1 расположен на основании, которое экстенсивно взаимодействует с субъединицей слияния (GP2), в которой находится петля слияния (26). Эндосомальные протеазы катепсин B (Cat B) и катепсин L (Cat L) необходимы для проникновения EBOV в цитоплазму и, следовательно, для инфицирования клеток-хозяев (10, 29, 45). Cat B и Cat L расщепляют GP внутри неструктурированной  13-  14 петли GP1, генерируя промежуточное соединение 20-kDa, а затем ключевое ядро 19-kDa (17, 25, 45). Процессинг, по-видимому, проходит через промежуточное соединение ϳ 50 кДа, которое напоминает GP ⌬.Во время этих массовых событий расщепления GP2 полностью защищен и остается дисульфидно связанным с GP1. Процессинг до GP 19 кДа удаляет муциноподобный домен, гликановый кэп и внешнюю β-цепь ( 14) головного домена (GP1). Район связывания рецептора (RBR) и основание GP1, а также весь GP2 остаются интактными. Мы определили расщепление GP1 катепсином как стадию праймирования, поскольку мы предположили, что GP2 остается зажатым в ядре 19 кДа (45). Доказательства в поддержку этого предложения включают наблюдение, что проникновение псевдовирионов, несущих 19-кДа GP, блокируется как бафиломицином, так и E64(d) (45, 54), предполагая, что низкий pH и чувствительный к E64 процесс необходимы для индукции 19- kDa GP для обеспечения слияния и проникновения.Первая не исключающая друг друга гипотеза (45), объясняющая эти находки, заключается в том, что праймирование ядра 19-kDa усиливает реакцию GP на триггер(ы) слияния. Во-вторых, расщепление до 19 кДа усиливает способность GP связываться с эндосомальным рецептором (29). Большинство вирусных слитых белков находятся на соответствующих поверхностях вирионов в метастабильном состоянии. Воздействие триггера слияния инициирует конформационные изменения, которые переводят слитый белок в более низкое энергетическое состояние и способствуют слиянию. Для всех вирусных слитых белков важным этапом запуска является экспозиция и изменение положения слитого пептида (или слитой петли) таким образом, чтобы он мог связываться с мембраной-мишенью.Только когда слитый пептид (петля) взаимодействует с мембраной-мишенью, последующая сворачивание гликопротеина может привести к слиянию бислоя вируса и бислоя-мишени (22, 53). В настоящем исследовании мы впервые разработали анализ, с помощью которого можно измерить основной ранний этап запуска слияния для EBOV GP: зависимое от петли слияния связывание с мембранами-мишенями. Затем мы использовали этот анализ для изучения роли расщепления катепсина и низкого pH в запуске EBOV GP. Наконец, мы получили доказательства того, что умеренное снижение может вызвать GP 19 кДа.Наши результаты предоставляют первое свидетельство триггерного конформационного изменения в EBOV GP, а также сильную поддержку гипотезы о том, что основная роль расщепления катепсином, особенно в отношении 19-kDa GP, заключается в усилении EBOV GP для запуска слияния.
Воздействие триггера слияния инициирует конформационные изменения, которые переводят слитый белок в более низкое энергетическое состояние и способствуют слиянию. Для всех вирусных слитых белков важным этапом запуска является экспозиция и изменение положения слитого пептида (или слитой петли) таким образом, чтобы он мог связываться с мембраной-мишенью.Только когда слитый пептид (петля) взаимодействует с мембраной-мишенью, последующая сворачивание гликопротеина может привести к слиянию бислоя вируса и бислоя-мишени (22, 53). В настоящем исследовании мы впервые разработали анализ, с помощью которого можно измерить основной ранний этап запуска слияния для EBOV GP: зависимое от петли слияния связывание с мембранами-мишенями. Затем мы использовали этот анализ для изучения роли расщепления катепсина и низкого pH в запуске EBOV GP. Наконец, мы получили доказательства того, что умеренное снижение может вызвать GP 19 кДа.Наши результаты предоставляют первое свидетельство триггерного конформационного изменения в EBOV GP, а также сильную поддержку гипотезы о том, что основная роль расщепления катепсином, особенно в отношении 19-kDa GP, заключается в усилении EBOV GP для запуска слияния. Кроме того, наши результаты дают новое представление о потенциальных механизмах запуска термоядерного синтеза. липосомы. Ранее мы показали (45), что инфекция псевдовирионами, несущими 19-kDa GP, продуцируемыми путем расщепления псевдовирионов GP либо Cat B плюс Cat L, либо термолизином, сильно ингибируется бафиломицином, агентом, повышающим pH эндосом.Это открытие свидетельствует о том, что слитая субъединица (GP2) в 19-кДа GP все еще зажата, т. е. ее петля слияния не подвергается воздействию, и, следовательно, 19-кДа GP находится в метастабильном состоянии. В качестве первой проверки этой гипотезы мы экспрессировали и очистили эктодомен EBOV GP, лишенный муциноподобного домена (GP ⌬ Ecto) (31), расщепили его термолизином, чтобы получить 19-кДа GP Ecto, и спросили, можно ли добавить тепло или мочевина могут индуцировать связывание мембраны-мишени. Этот тип экспериментов подтвердил метастабильность многих слитых белков вирусов класса I, в том числе белков вируса гриппа (9), PIV5 (11), вируса саркомы птиц и лейкоза (ASLV) (37, 49) и вируса мышиного лейкоза Молони ( МоМЛВ) (51).
Кроме того, наши результаты дают новое представление о потенциальных механизмах запуска термоядерного синтеза. липосомы. Ранее мы показали (45), что инфекция псевдовирионами, несущими 19-kDa GP, продуцируемыми путем расщепления псевдовирионов GP либо Cat B плюс Cat L, либо термолизином, сильно ингибируется бафиломицином, агентом, повышающим pH эндосом.Это открытие свидетельствует о том, что слитая субъединица (GP2) в 19-кДа GP все еще зажата, т. е. ее петля слияния не подвергается воздействию, и, следовательно, 19-кДа GP находится в метастабильном состоянии. В качестве первой проверки этой гипотезы мы экспрессировали и очистили эктодомен EBOV GP, лишенный муциноподобного домена (GP ⌬ Ecto) (31), расщепили его термолизином, чтобы получить 19-кДа GP Ecto, и спросили, можно ли добавить тепло или мочевина могут индуцировать связывание мембраны-мишени. Этот тип экспериментов подтвердил метастабильность многих слитых белков вирусов класса I, в том числе белков вируса гриппа (9), PIV5 (11), вируса саркомы птиц и лейкоза (ASLV) (37, 49) и вируса мышиного лейкоза Молони ( МоМЛВ) (51). Вышеупомянутые исследования отслеживали ассоциацию эктодомена слитого белка с липосомами-мишенями с использованием градиентов флотации сахарозы (7, 16, 20, 48). Чтобы увеличить производительность анализа, мы модифицировали метод с использованием липосом, содержащих биотинилированный липид (34). Если эктодомен слитого белка связывается с биотинилированными липосомами, он останется связанным с липосомами после инкубации с магнитными шариками, связанными со стрептавидином. Сначала мы протестировали анализ с хорошо охарактеризованным фрагментом гемагглютинина гриппа (BHA), активируемым при низких значениях pH и высвобождаемым бромелаином.Когда BHA подвергали воздействию pH 5 в течение 5 минут при 37°C, большая часть была притянута магнитными шариками стрептавидина (благодаря связыванию с целевыми биотинилированными липосомами), тогда как только следовые количества были связаны с шариками в образцах, обработанных при pH 7. в течение 5 мин при 37°С (данные не представлены). Такое поведение согласуется с результатами, полученными с использованием градиентов флотации липосом (16, 48). Затем мы использовали анализ связывания липосом с магнитными шариками, чтобы выяснить, может ли повышенная температура или мочевина вызвать связывание 19-kDa GP Ecto с липосомами.Мы проводили анализы как при нейтральном, так и при низком рН, поскольку считается, что GP-опосредованное слияние происходит в поздних эндосомах (10, 43, 45), которые имеют рН ϳ 5, и поскольку низкий рН необходим для проникновения псевдовирионов, несущих ГП 19 кДа (45). Как видно на фиг. 1А, обработка 19-кДа GP Ecto мочевиной при pH 5,0, но не при нейтральном pH, индуцировала связывание 19-kDa GP Ecto с липосомами. Пик связывания наблюдался при воздействии на GP Ecto 19 кДа 1,5 М мочевины. Повышенная температура также индуцировала связывание 19-кДа GP Ecto с липосомами (рис.1Б). При рН 5,0 низкий уровень связывания наблюдался при 37°С, а максимальный уровень связывания наблюдался при температуре от 50°С до 60°С. Никакого связывания не наблюдалось при рН 7,5 при 37°С, а при 50°С обнаруживалось лишь следовое количество.
Вышеупомянутые исследования отслеживали ассоциацию эктодомена слитого белка с липосомами-мишенями с использованием градиентов флотации сахарозы (7, 16, 20, 48). Чтобы увеличить производительность анализа, мы модифицировали метод с использованием липосом, содержащих биотинилированный липид (34). Если эктодомен слитого белка связывается с биотинилированными липосомами, он останется связанным с липосомами после инкубации с магнитными шариками, связанными со стрептавидином. Сначала мы протестировали анализ с хорошо охарактеризованным фрагментом гемагглютинина гриппа (BHA), активируемым при низких значениях pH и высвобождаемым бромелаином.Когда BHA подвергали воздействию pH 5 в течение 5 минут при 37°C, большая часть была притянута магнитными шариками стрептавидина (благодаря связыванию с целевыми биотинилированными липосомами), тогда как только следовые количества были связаны с шариками в образцах, обработанных при pH 7. в течение 5 мин при 37°С (данные не представлены). Такое поведение согласуется с результатами, полученными с использованием градиентов флотации липосом (16, 48). Затем мы использовали анализ связывания липосом с магнитными шариками, чтобы выяснить, может ли повышенная температура или мочевина вызвать связывание 19-kDa GP Ecto с липосомами.Мы проводили анализы как при нейтральном, так и при низком рН, поскольку считается, что GP-опосредованное слияние происходит в поздних эндосомах (10, 43, 45), которые имеют рН ϳ 5, и поскольку низкий рН необходим для проникновения псевдовирионов, несущих ГП 19 кДа (45). Как видно на фиг. 1А, обработка 19-кДа GP Ecto мочевиной при pH 5,0, но не при нейтральном pH, индуцировала связывание 19-kDa GP Ecto с липосомами. Пик связывания наблюдался при воздействии на GP Ecto 19 кДа 1,5 М мочевины. Повышенная температура также индуцировала связывание 19-кДа GP Ecto с липосомами (рис.1Б). При рН 5,0 низкий уровень связывания наблюдался при 37°С, а максимальный уровень связывания наблюдался при температуре от 50°С до 60°С. Никакого связывания не наблюдалось при рН 7,5 при 37°С, а при 50°С обнаруживалось лишь следовое количество. Наши стандартные липосомы (см. Материалы и методы) состоят из PC-PE-BtPE-Sph-Chol (1,0:0,9:0,1:1,0:1,5). В образцах, нагретых до 50°C при pH 5, 19-кДа GP Ecto одинаково хорошо связывался с липосомами, в которых половина (небиотинилированного) PE была заменена либо ПОПГ [1-пальмитоил-2-олеоил-sn-глицеро-3 -фосфо-(1=-rac глицерин)], LBPA (лизобисфосфатидная кислота) или POPS (1-пальмитоил-2-олеоил-sn-глицеро-3-фосфо-L-серин) (данные не показаны).На рис. 2 представлен более подробный анализ температурной зависимости связывания липосом. При рН 5 ассоциация практически не наблюдалась при 22°С, резкий рост наблюдался между 37°С и 45°С, максимальная ассоциация наблюдалась при ϳ 50°С (рис. 2А). Температура влияла как на скорость, так и на степень связывания 19-кДа GP Ecto с липосомами (рис. 2В). При 50°С связывание быстро возрастало, достигая кажущегося максимума через 2 мин; при 42°С и 37°С связывание с липосомами происходило прогрессивно медленнее и меньше…
Наши стандартные липосомы (см. Материалы и методы) состоят из PC-PE-BtPE-Sph-Chol (1,0:0,9:0,1:1,0:1,5). В образцах, нагретых до 50°C при pH 5, 19-кДа GP Ecto одинаково хорошо связывался с липосомами, в которых половина (небиотинилированного) PE была заменена либо ПОПГ [1-пальмитоил-2-олеоил-sn-глицеро-3 -фосфо-(1=-rac глицерин)], LBPA (лизобисфосфатидная кислота) или POPS (1-пальмитоил-2-олеоил-sn-глицеро-3-фосфо-L-серин) (данные не показаны).На рис. 2 представлен более подробный анализ температурной зависимости связывания липосом. При рН 5 ассоциация практически не наблюдалась при 22°С, резкий рост наблюдался между 37°С и 45°С, максимальная ассоциация наблюдалась при ϳ 50°С (рис. 2А). Температура влияла как на скорость, так и на степень связывания 19-кДа GP Ecto с липосомами (рис. 2В). При 50°С связывание быстро возрастало, достигая кажущегося максимума через 2 мин; при 42°С и 37°С связывание с липосомами происходило прогрессивно медленнее и меньше…
экто-5’NT/CD73 при метастазировании как потенциальный прогностический фактор
Abstract
Медуллобластома (МБ) является наиболее частой злокачественной опухолью головного мозга у детей и возникает преимущественно в мозжечке. Важные внутриклеточные сигнальные молекулы, такие как присутствующие в путях Sonic Hedgehog и Wnt, участвуют в его развитии и могут также использоваться для определения степени опухоли и прогноза. Эктонуклеотидазы, особенно экто-5’NT/CD73, являются важными ферментами злокачественного процесса различных типов опухолей, регулирующими внеклеточный уровень АТФ и аденозина.Здесь мы исследовали активность эктонуклеотидаз в трех линиях злокачественных клеток человека: Daoy и ONS76, представляющих первичный MB, и клеточной линии D283, полученной из метастатического MB. Все клеточные линии секретировали АТФ во внеклеточную среду, но плохо гидролизовали этот нуклеотид, что согласуется с низкой экспрессией и активностью пирофосфат/фосфодиэстеразы, НТФДаз и щелочной фосфатазы. Анализ гидролиза AMP показал, что Daoy и ONS76 полностью гидролизовали AMP с параллельным образованием аденозина (Daoy) и накоплением инозина (ONS76).С другой стороны, клеточная линия D283 не гидролизовала АМФ. Кроме того, первичные опухолевые клетки MB, Daoy и ONS76, экспрессируют экто-5’NT/CD73, в то время как D283, представляющий метастатическую опухоль, показал слабую экспрессию этого фермента, в то время как экто-аденозиндезаминаза показала более высокую экспрессию в D283 по сравнению с Daoy и ONS76. клетки. Ядерный бета-катенин был предложен в качестве маркера прогноза MB. Кроме того, он может способствовать экспрессии экто-5’NT/CD73 и подавлению аденозиндезаминазы. Было замечено, что Daoy и ONS76 показали большую ядерную иммунореактивность к бета-катенину, чем D283, который проявлял в основном цитоплазматическую иммунореактивность.Таким образом, отсутствие экто-5’NT/CD73 в клеточной линии D283, метастатическом фенотипе MB, предполагает, что высокие уровни экспрессии этой эктонуклеотидазы могут быть связаны с плохим прогнозом у пациентов с MB.
Образец цитирования: Капеллари А.Р., Рокенбах Л., Дитрих Ф., Кларимундо В., Глейзер Т., Браганхол Э. и др. (2012) Характеристика эктонуклеотидаз в клеточных линиях медуллобластомы человека: экто-5’NT/CD73 в метастазах как потенциальный прогностический фактор. ПЛОС ОДИН 7(10): е47468.https://doi.org/10.1371/journal.pone.0047468
Редактор: Тобиас Экл, Университет Колорадо Денвер, Соединенные Штаты Америки
Поступила в редакцию: 12 июля 2012 г. ; Принято: 11 сентября 2012 г.; Опубликовано: 18 октября 2012 г.
; Принято: 11 сентября 2012 г.; Опубликовано: 18 октября 2012 г.
Авторское право: © Cappellari et al. Это статья с открытым доступом, распространяемая в соответствии с лицензией Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания оригинального автора и источника.
Финансирование: Эта работа была поддержана бразильскими финансирующими агентствами: CNPq (Proc. 303016/2009-4), FIPEHCPA (Proc. 10-01320), FAPESP (Henning Ulrich). A. Cappellari был получателем стипендии Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). Спонсоры не участвовали в разработке исследования, сборе и анализе данных, решении о публикации или подготовке рукописи.
Конкурирующие интересы: Хеннинг Ульрих является членом редакционной коллегии PLoS ONE, и мы подтверждаем, что это не меняет нашей приверженности всем политикам PLoS ONE в отношении обмена данными и материалами, как подробно описано в нашем онлайн-руководстве для авторов.
Введение
Медуллобластома (МБ) является наиболее распространенной злокачественной опухолью головного мозга у детей. Он поражает преимущественно мальчиков в возрасте около 9 лет, при этом в нескольких случаях сообщается о MB среди взрослых старше 20 лет. МБ возникает в основном в мозжечке и задней черепной ямке, имея возможность мигрировать в другие структуры головного мозга. Эта опухоль дает экстраневральные метастазы в 7,1% случаев, что связано с ее неблагоприятным прогнозом [1], [2]. Всемирная организация здравоохранения (ВОЗ) классифицировала МБ как опухоль IV степени, наиболее злокачественную [3].Эта опухоль подразделяется на подмножества, включая десмопластическую/узловую, анапластическую, МБ с обширными узлами и крупноклеточную МБ [3], [4]. Sonic Hedgehog, C-MYC, NOTCH и Wnt индуцируют основные внутриклеточные сигнальные пути, участвующие в развитии MB, и связаны с различными подмножествами MB и степенью опухоли [4]–[6]. Мутации в нижестоящих эффекторах пути Wnt, включая белок APC и β-catenin, присутствующие в спорадических MB, предполагают, что аберрантная передача сигналов пути участвует в развитии заболевания. Кроме того, недавние исследования показали, что ядерная или цитоплазматическая иммунореактивность β-катенина может быть связана с хорошим или плохим прогнозом у пациентов с МБ соответственно [7].
Известно множество факторов, влияющих на прогрессирование злокачественной опухоли. Среди них можно выделить пуринергическую систему, идентифицированную во многих типах опухолей [8]–[11]. АТФ представляет собой нуклеотид, который может высвобождаться во внеклеточную среду в ответ на физиологические или патологические состояния и выполнять различные функции, активируя P2X, определяемый как класс ионотропных или лиганд-управляемых рецепторов, и рецепторы P2Y, классифицируемые как метаботропные рецепторы, связанные с G-белком. 12], [13].
Передача сигналов АТФ модулируется общим классом эффективных ферментов, называемых эктонуклеотидазами. Они подразделяются на E-NTPDases (эктонуклеотидтрифосфатдифосфогидролазы), E-NPP (эктонуклеотидпирофосфат/фосфодиэстераза), Ecto-5’NT/CD73 (экто-5’нуклеотидаза/CD73) и ALP (экто-щелочная фосфатаза). ) [14]. NTPDases1, 2, 3 и 8 являются эктоферментами, которые различаются по своему субстратному предпочтению между АТФ и АДФ, а также по их распределению в тканях. Внутриклеточно экспрессируемые NTPDase5 и 6 секретируются во внеклеточную среду при стимуляции, тогда как NTPDases4 и 7 являются исключительно внутриклеточными ферментами, локализованными в просвете органелл.Семейство E-NPP состоит из семи эктоферментов, но только NPP1, 2 и 3 участвуют в пуринергической передаче сигналов из-за их способности гидролизовать широкий спектр фосфатных субстратов, таких как превращение АТФ в AMP и PPi и АДФ в AMP и Пи [14]–[16].
) [14]. NTPDases1, 2, 3 и 8 являются эктоферментами, которые различаются по своему субстратному предпочтению между АТФ и АДФ, а также по их распределению в тканях. Внутриклеточно экспрессируемые NTPDase5 и 6 секретируются во внеклеточную среду при стимуляции, тогда как NTPDases4 и 7 являются исключительно внутриклеточными ферментами, локализованными в просвете органелл.Семейство E-NPP состоит из семи эктоферментов, но только NPP1, 2 и 3 участвуют в пуринергической передаче сигналов из-за их способности гидролизовать широкий спектр фосфатных субстратов, таких как превращение АТФ в AMP и PPi и АДФ в AMP и Пи [14]–[16].
Ecto-5’NT/CD73 представляет собой широко распространенный фермент, связанный с плазматической мембраной гликозил-фосфатидилинозитол-липидным якорем. Его основная функция заключается в гидролизе АМФ в аденозин во внеклеточной среде. Кроме того, этот фермент выполняет неферментативные функции, участвуя во взаимодействиях между клетками и клетками и матриксом и стимулируя внутриклеточные сигнальные пути [15], [17], [18]. Эктощелочные фосфатазы (ЩФ) состоят из четырех ферментов, широко распространенных в организме человека. Они гидролизуют широкий спектр субстратов, включая АТФ, АДФ, АМФ и PPi [19]. Однако ЩФ составляют важную группу ферментов, которые главным образом гидролизуют АМФ, участвуя во внеклеточном метаболизме нуклеотидов в качестве продуцента аденозина [20], [21]. Наконец, доступность внеклеточного аденозина контролируется переносчиками аденозина, расположенными в плазматической мембране, и/или действием экто-аденозиндезаминазы (ADA), продуцирующей инозин [22].
Эктощелочные фосфатазы (ЩФ) состоят из четырех ферментов, широко распространенных в организме человека. Они гидролизуют широкий спектр субстратов, включая АТФ, АДФ, АМФ и PPi [19]. Однако ЩФ составляют важную группу ферментов, которые главным образом гидролизуют АМФ, участвуя во внеклеточном метаболизме нуклеотидов в качестве продуцента аденозина [20], [21]. Наконец, доступность внеклеточного аденозина контролируется переносчиками аденозина, расположенными в плазматической мембране, и/или действием экто-аденозиндезаминазы (ADA), продуцирующей инозин [22].
В последние годы во многих сообщениях показано участие пуринергической системы в опухолевой прогрессии с акцентом на участие эктонуклеотидаз в патогенезе этого процесса. Наши предыдущие результаты показывают, что клеточные линии с высокой степенью злокачественности демонстрируют повышенный уровень экто-5’NT/CD73 и не имеют активности E-NTPDase [23]. Отсутствие E-NTPDases приводит к накоплению АТФ [24], а присутствие экто-5’NT/CD73 способствует выработке аденозина из AMP, который высвобождается при гибели нормальной ткани, окружающей опухоль [25]. И АТФ, и аденозин in vitro стимулировали пролиферацию клеток глиомы [26]. Кроме того, глиомы демонстрируют положительную корреляцию между увеличением слияния клеток с повышенной экспрессией экто-5’NT/CD73, что свидетельствует об участии этого фермента в прогрессировании этой опухоли [9].
И АТФ, и аденозин in vitro стимулировали пролиферацию клеток глиомы [26]. Кроме того, глиомы демонстрируют положительную корреляцию между увеличением слияния клеток с повышенной экспрессией экто-5’NT/CD73, что свидетельствует об участии этого фермента в прогрессировании этой опухоли [9].
Линии клеток MB, используемые в этом исследовании, считаются важными моделями для оценки прогрессирования и злокачественности [27]–[30]. Клеточные линии Daoy и ONS76 считаются репрезентативными для первичного MB.С другой стороны, клеточная линия D283 представляет собой MB, образующий метастазы в брюшине, из которого была получена эта клеточная линия. В связи с этим мы предложили охарактеризовать профиль экспрессии и активности эктонуклеотидаз в клеточных линиях МБ. Здесь мы сообщаем, что клеточные линии Daoy, ONS76 и D283 различаются по уровням экспрессии эктонуклеотидазы и ферментативной активности. Мы также показали, что вариации экспрессии экто-5’NT/CD73 и ADA могут быть связаны с соответствующими уровнями иммунореактивности β-катенина, что свидетельствует о важности этих ферментов в качестве маркеров прогноза.
Материалы и методы
Реагенты
Среда для культивирования клеток, пенициллин/стрептомицин, раствор трипсина/ЭДТА и фетальная телячья сыворотка были приобретены у Gibco (Gibco BRL, Карлсбад, Калифорния, США). АТФ, АДФ и все монофосфатные нуклеозиды, аденозин, натриевая соль тимидин-5′-монофосфата ρ-нитрофенилового эфира, 4-нитрофенол, β -глицерофосфат, D- глюкозо-6-фосфат натриевая соль и левамизол были получены от Sigma. (Сигма Олдрич Корпорейшн, Ст.Луис, США). Все другие используемые химические вещества были аналитической чистоты.
Культура клеток
Клеточные линии Daoy и ONS76 (представитель первичной опухоли человека) и D283 (представитель метастатического MB человека) были любезно предоставлены Исследовательской лабораторией рака и Институтом детского рака (HCPA и ICI-RS, Риу-Гранди-ду-Сул, Бразилия). Клеточные линии Daoy и D283 были первоначально получены из ATCC (Американская коллекция типовых культур), тогда как клеточная линия ONS76 была первоначально получена из JCRB (Японская коллекция исследовательских биоресурсов). Клетки выращивали и поддерживали в модифицированной по Дульбекко среде Игле (DMEM) с низким содержанием глюкозы, содержащей 0,5 ед/мл антибиотиков пенициллин/стрептомицин и дополненной 10% фетальной бычьей сывороткой (FBS). Клетки хранили при 37°С в инкубаторе с минимальной относительной влажностью 95% и 5% СО 2 в воздухе.
Клетки выращивали и поддерживали в модифицированной по Дульбекко среде Игле (DMEM) с низким содержанием глюкозы, содержащей 0,5 ед/мл антибиотиков пенициллин/стрептомицин и дополненной 10% фетальной бычьей сывороткой (FBS). Клетки хранили при 37°С в инкубаторе с минимальной относительной влажностью 95% и 5% СО 2 в воздухе.
Анализ нуклеотидов и нуклеозидов методом ВЭЖХ
Клетки высевали с плотностью 10 90 109 4 90 110 клеток/лунку в 24-многолуночные планшеты. После достижения слияния клетки трижды промывали инкубационной средой, содержащей 2 мМ MgCl 2 , 120 мМ NaCl, 5 мМ KCl, 10 мМ глюкозы, 20 мМ Hepes, pH 7.4. Реакцию начинали добавлением AMP в конечной концентрации 100 мкМ в 250 мкл при 37°C. После разного времени инкубации (0, 5, 10, 20, 30, 60, 90 мин) отбирали надосадочную жидкость и выдерживали на льду. Для определения базальной секреции нуклеотидов клетки инкубировали в течение 90 мин со средой инкубации без субстрата. Собранные супернатанты центрифугировали в течение 30 минут при 16000×g и 4°C, аликвоты по 40 мкл анализировали с помощью системы ВЭЖХ с обращенной фазой (Supelcosil LC-18, 25 см×4. 6 мм, Supelco) в жидкостном хроматографе Shimadzu Instruments (Shimadzu, Япония). Разделение проводили с использованием линейного градиента от 100 % растворителя А (60 мМ KH 2 PO 4 и 5 мМ тетрабутиламмонийфосфата, pH 6,0) до 100 % растворителя B (70 % 100 мМ KH 2 PO 4 и 5 мМ тетрабутиламмонийфосфата, pH 6,0 в 30% метаноле). Количество пуринов определяли по поглощению при 254 нм, и все пики идентифицировали по времени удерживания и количественно определяли путем сравнения со стандартами.Концентрации пуринов выражены в нмоль продукта/мг белка (среднее значение ± стандартное отклонение). Все анализы проводили в трех экземплярах, а контроли, используемые для коррекции неферментативного гидролиза нуклеотидов, проводили путем измерения пиков, присутствующих в той же реакционной среде в отсутствие клеток.
ПЦР с обратной транскриптазой и ПЦР в реальном времени
Тотальную РНК из культур клеточной линии MB выделяли с помощью реагента Trizol LS в соответствии с инструкциями производителя. Образцы РНК обрабатывали ДНКазой (амплификация набора дезоксирибонуклеазы I — Sigma-Aldrich Brasil Ltda) в течение 10 мин при 37°C и образцы инкубировали в течение 10 мин при 65°C для остановки реакции.кДНК синтезировали с помощью обратной транскриптазы RevertAid (Fermentas – Life Sciences) из 3 мкг тотальной РНК в конечном объеме 20 мкл с праймером oligo dT в соответствии с инструкциями производителя. В последовательности реакции кДНК проводили в течение 1 часа при 42°С и останавливали нагреванием до 70°С. Для ОТ-ПЦР использовали реакционные смеси общим объемом 25 мкл, которые включали 1 мкл реакции ОТ, 2,5 мкл 10× Taq ДНК-полимеразного буфера, 1,5 мкл 25 мМ MgCl 2 , 0,5 мкл 10 мкМ смеси dNTP. , 1 мкл каждого конкретного праймера в концентрации 10 мкМ (таблица 1) и 1 мкл фермента ДНК-полимеразы Taq (Fermentas – Life Sciences).Условия циклирования ПЦР были следующими: 2 минуты при 95°С, 30 секунд при 95°С, 30 секунд при температуре отжига и 1 минута при 72°С. Все реакции ПЦР проводили в течение 35 циклов и включали заключительную 10-минутную доработку при 72°С. Десять микролитров реакции ПЦР анализировали на 2,0% агарозном геле, содержащем бромистый этидий, и визуализировали в ультрафиолетовом свете. В качестве контроля синтеза кДНК проводили ПЦР с глицеральдегид-3-фосфатдегидрогеназой (GAPDH). Плазмиды, содержащие последовательности NTPDase1, NTPDase2, NTPDase3 и экто-5′-нуклеотидазы/CD73, использовали в качестве положительного контроля экспрессии этих ферментов; кДНК Т24 использовали в качестве положительного контроля экспрессии гена NTPDase5.Для анализа экспрессии генов NPP1, 2 и 3, ALP и ADA использовали смесь кДНК клеточных линий человека. Отрицательные контроли выполняли путем замены матриц на дистиллированную воду, не содержащую ДНКазы/РНКазы, в каждой реакции ПЦР.
Образцы РНК обрабатывали ДНКазой (амплификация набора дезоксирибонуклеазы I — Sigma-Aldrich Brasil Ltda) в течение 10 мин при 37°C и образцы инкубировали в течение 10 мин при 65°C для остановки реакции.кДНК синтезировали с помощью обратной транскриптазы RevertAid (Fermentas – Life Sciences) из 3 мкг тотальной РНК в конечном объеме 20 мкл с праймером oligo dT в соответствии с инструкциями производителя. В последовательности реакции кДНК проводили в течение 1 часа при 42°С и останавливали нагреванием до 70°С. Для ОТ-ПЦР использовали реакционные смеси общим объемом 25 мкл, которые включали 1 мкл реакции ОТ, 2,5 мкл 10× Taq ДНК-полимеразного буфера, 1,5 мкл 25 мМ MgCl 2 , 0,5 мкл 10 мкМ смеси dNTP. , 1 мкл каждого конкретного праймера в концентрации 10 мкМ (таблица 1) и 1 мкл фермента ДНК-полимеразы Taq (Fermentas – Life Sciences).Условия циклирования ПЦР были следующими: 2 минуты при 95°С, 30 секунд при 95°С, 30 секунд при температуре отжига и 1 минута при 72°С. Все реакции ПЦР проводили в течение 35 циклов и включали заключительную 10-минутную доработку при 72°С. Десять микролитров реакции ПЦР анализировали на 2,0% агарозном геле, содержащем бромистый этидий, и визуализировали в ультрафиолетовом свете. В качестве контроля синтеза кДНК проводили ПЦР с глицеральдегид-3-фосфатдегидрогеназой (GAPDH). Плазмиды, содержащие последовательности NTPDase1, NTPDase2, NTPDase3 и экто-5′-нуклеотидазы/CD73, использовали в качестве положительного контроля экспрессии этих ферментов; кДНК Т24 использовали в качестве положительного контроля экспрессии гена NTPDase5.Для анализа экспрессии генов NPP1, 2 и 3, ALP и ADA использовали смесь кДНК клеточных линий человека. Отрицательные контроли выполняли путем замены матриц на дистиллированную воду, не содержащую ДНКазы/РНКазы, в каждой реакции ПЦР.
ПЦР-анализ в реальном времени выполняли на приборе ABI Step One Plus (Applied Biosystems, Фостер-Сити, Калифорния) с использованием системы амплификации Sybr Green. Каждую реакцию проводили в конечном объеме 25 мкл, содержащем 1 мкл кДНК, обратно транскрибированной из 3 мкг РНК, 12. 5 мкл Sybr Green Universal PCR Master Mix (Applied Biosystems, Фостер-Сити, Калифорния) и по 0,3 мкл каждого прямого и обратного праймеров (10 мкМ) (таблица 1). Реакции ПЦР в реальном времени проводили с использованием следующих температурных протоколов: 95°С в течение 10 минут, 40 циклов при 95°С в течение 15 секунд каждый и 60°С в течение 1 минуты. Поскольку эффективность всех реакций была >95%, параметр ΔΔCt использовали для определения данных относительной экспрессии, принимая экспрессию гена GAPDH в качестве эндогенного контроля для нормализации.Стандартные кривые измеряли для каждого набора праймеров и образца кДНК, чтобы проверить эффективность реакции.
5 мкл Sybr Green Universal PCR Master Mix (Applied Biosystems, Фостер-Сити, Калифорния) и по 0,3 мкл каждого прямого и обратного праймеров (10 мкМ) (таблица 1). Реакции ПЦР в реальном времени проводили с использованием следующих температурных протоколов: 95°С в течение 10 минут, 40 циклов при 95°С в течение 15 секунд каждый и 60°С в течение 1 минуты. Поскольку эффективность всех реакций была >95%, параметр ΔΔCt использовали для определения данных относительной экспрессии, принимая экспрессию гена GAPDH в качестве эндогенного контроля для нормализации.Стандартные кривые измеряли для каждого набора праймеров и образца кДНК, чтобы проверить эффективность реакции.
Анализ активности эктонуклеотидной пирофосфатазы/фосфодиэстеразы
Активность E-NPP оценивали с использованием 0,5 мМ тимидин-5′-монофосфата ρ-нитрофенила (5′-TMP-p-Nph), маркера субстрата, который обычно используется для анализа in vitro этих ферментов, как описано ранее. [31]. Клетки инкубировали в реакционной среде, содержащей 50 мМ трис-HCl-буфера (pH 7. 4 или 8.9), 5 мМ KCl, 120 мМ NaCl, 0,5 мМ CaCl 2 , 60 мМ глюкозы и 0,5 мМ 5′-TMP-p-Nph. Реакцию останавливали добавлением 200 мкл 0,2 М NaOH. Количество высвобожденного ρ-нитрофенола определяли по изменению оптической плотности (OD) при 400 нм в ридере для микропланшетов. Удельную активность выражали в нмоль ρ-нитрофенола/мг белка.
Ферментативный анализ на эктонуклеотидазу
Для определения гидролиза нуклеотидов 24-луночные планшеты, содержащие опухолевые клетки Daoy, ONS76 и D283 MB, трижды промывали бесфосфатной инкубационной средой в отсутствие нуклеотидов.Реакцию начинали добавлением 200 мкл инкубационной среды, содержащей 2 мМ CaCl 2 , 120 мМ NaCl, 5 мМ KCl, 10 мМ глюкозы, 20 мМ HEPES (рН 7,4) и 1 мМ АТФ или АДФ при 37°С. . Экспериментальные условия были такими же, как описано выше для количественного определения гидролиза монофосфонуклеозидов и сложных эфиров фосфорной кислоты, за исключением того, что вместо CaCl 2 использовали 2 мМ MgCl 2 . Тестируемые субстраты включали AMP, CMP, GMP, IMP, UMP, неорганический пирофосфат (PPi), глюкозо-6-фосфат (Glu-6P) и β-глицерофосфат (β-Gly).Концентрации субстрата и время инкубации выбирали таким образом, чтобы обеспечить линейность реакции. Реакцию останавливали, отбирая аликвоту инкубационной среды и перенося ее в предварительно охлажденную пробирку, содержащую ТХУ (10% мас./об.). Выделившийся неорганический фосфат (Pi) измеряли методом малахитовой зелени [32], используя KH -2- PO -4- в качестве эталона Pi. Контроль для определения неферментативного высвобождения Pi проводили путем инкубации клеток в отсутствие субстрата в отсутствие клеток.Все образцы были выполнены в трехкратной повторности. Удельную активность выражали в нмоль высвобожденного Pi/мин/мг белка (нмоль Pi/мин/мг).
Определение белка
Для количественного определения концентрации белка клетки высушивали, солюбилизировали 100 мкл 1 н. NaOH и замораживали в течение ночи. Концентрацию белка измеряли методом кумасси синего [33] с использованием в качестве стандарта бычьего сывороточного альбумина (БСА).
Анализ проточной цитометрии
Для анализа методом проточной цитометрии линии раковых клеток Daoy, ONS76 и D283 MB содержались в культуральных флаконах до тех пор, пока они не достигали слияния.Затем клетки обрабатывали трипсином, диссоциировали и немедленно подсчитывали в гемоцитометре. Затем 1×10 90 109 6 90 110 клеток центрифугировали в течение 5 мин при 400 g и дважды промывали фосфатно-солевым буфером (PBS) с добавлением 1% фетальной телячьей сыворотки (FCS). Осадок суспендировали и инкубировали в течение 1 ч с очищенным мышиным антителом против человеческого CD73 (BD Pharmingen TM) (1×10) в течение 1 ч при 4°C. Параллельно такое же количество клеток инкубировали без антител. Последовательно все образцы промывали PBS плюс 1% FCS и центрифугировали.Осадки суспендировали и инкубировали в течение 1 ч с кроличьим антимышиным IgG Alexa fluor 555 (1∶100). После этого меченые клетки промывали PBS и немедленно анализировали с помощью проточной цитометрии (FACS Calibur, BD Biosciences, Mountain View, CA). Десять тысяч событий в клеточных воротах были собраны и дополнительно проанализированы с использованием программного обеспечения CellQuest (BD Biosciences) и программного обеспечения FCS Express 4.
Десять тысяч событий в клеточных воротах были собраны и дополнительно проанализированы с использованием программного обеспечения CellQuest (BD Biosciences) и программного обеспечения FCS Express 4.
Экстракция ядерных и цитозольных фракций и вестерн-блоттинг
Для приготовления клеточных лизатов клетки МБ трипсинизировали, центрифугировали 10 мин при 400 g, промывали PBS и повторно центрифугировали.Затем осадок растворяли в лизисном буфере (10 мМ Hepes, 1,5 мМ MgCl 2 , 10 мМ KCl, 0,1 мМ EDTA, 0,1 мМ EGTA, 0,5% NP40) плюс смесь ингибиторов протеаз (Fermentas – Life Sciences) и ингибиторов фосфатазы. (2 мМ ортованадата и 5 мМ фторида натрия), инкубировали 15 мин на льду, затем центрифугировали 5 мин при 13000 g и 4°С. Супернатант собирали и использовали в качестве растворимой цитозольной фракции. Осадок суспендировали в буфере ядерной фракции (20 мМ Hepes, 1,5 мМ MgCl 2 , 300 мМ NaCl, 0.25 мМ ЭДТА, 0,25 мМ ЭГТА, 25% глицерин), инкубировали 20 мин на льду при перемешивании и центрифугировали 20 мин при 13000 g и 4°С. Супернатант собирали как фракцию, обогащенную ядерным белком. Количественное определение белка измеряли методом кумасси синего [33] с бычьим сывороточным альбумином в качестве стандарта. Тридцать мкг белка в буфере для образцов разделяли с помощью SDS-PAGE на 10% полиакриламидном геле при постоянном напряжении 140 В. Затем белки переносили на нитроцеллюлозную мембрану (Thermo-scientific) во влажной системе в течение ночи при постоянной силе тока 20 В. мА.Для блокирования неспецифического связывания добавляли 5% БСА в TBS-T на 30 мин при перемешивании при комнатной температуре. Затем мембраны инкубировали с первичными антителами к фосфо-β-катенину (Ser33/37/Thr41) (Cell Signaling Tech.), HnRNPK (Santa Cruz Biotechnology, Санта-Крус, США) и GAPDH (Invitrogen, Life Technologies, Карлсбад, США). ) в течение ночи при 4°С. Затем мембраны промывали и зондировали соответствующими вторичными антителами в течение 1 ч при перемешивании при комнатной температуре. Первичные и вторичные антитела разводили в 5% BSA и TBS-T. Мембраны промывали в TBS-T и сканировали с помощью Typhoon™ — GE Healthcare. Полученные полосы подвергали денситометрическому анализу с помощью программного обеспечения ImageJ. Уровни фосфо-β-катенина нормализовали по сравнению с HnRNPK и GAPDH в ядерной и цитозольной фракциях соответственно.
Мембраны промывали в TBS-T и сканировали с помощью Typhoon™ — GE Healthcare. Полученные полосы подвергали денситометрическому анализу с помощью программного обеспечения ImageJ. Уровни фосфо-β-катенина нормализовали по сравнению с HnRNPK и GAPDH в ядерной и цитозольной фракциях соответственно.
Иммунофлуоресцентные исследования
Клетки фиксировали в течение 20 мин 4% параформальдегидом в PBS. Через 45 мин инкубации добавляли блокирующий раствор, содержащий 2% козьей сыворотки в сочетании с 0.1% Triton X100 (Sigma Aldrich Corp., Сент-Луис, США) в PBS. В этой последовательности клетки инкубировали в течение 2 ч с первичными антителами против фосфо-β-катенина (Ser33/37/Thr41) (Cell Signaling Tech.), разведенными в PBS плюс 0,1% Triton X-100 и 2% козьей сыворотки. После промывания PBS на 1 час добавляли вторичное антитело кролика Alexa fluor 555 против мышиного IgG (1∶100) с последующей 5-минутной инкубацией с DAPI (4′-6-диамино-2-фенилиндол). Клетки промывали PBS, помещали на предметные стекла с D. P.X. (Сигма Олдрич Корп., Сент-Луис, США) и исследовали с помощью инвертированного флуоресцентного микроскопа (Nikon Eclipse TE 300).
P.X. (Сигма Олдрич Корп., Сент-Луис, США) и исследовали с помощью инвертированного флуоресцентного микроскопа (Nikon Eclipse TE 300).
Рис. 1. Базальная секреция нуклеотидов клеточными линиями MB. Клеточные линии
MB инкубировали в течение 90 минут с инкубационной средой в отсутствие субстрата, как описано в разделе «Материалы и методы». Концентрации внеклеточных нуклеотидов/нуклеозидов (АТФ, АДФ, АМФ, АДО и INO) измеряли с помощью ВЭЖХ, и концентрации выражали как мкмоль продукта/мг белка (среднее ± стандартное отклонение). Эксперименты проводили трижды в трех повторностях, и полученные данные анализировали на статистическую релевантность с использованием теста двухфакторного дисперсионного анализа с последующим апостериорным тестом Бонферрони.(***) p<0,001 и (*) p<0,05 по сравнению с контрольными данными.
https://doi.org/10.1371/journal.pone.0047468.g001
Рисунок 2. Анализ экспрессии эктонуклеотидаз в клеточных линиях MB.
Суммарная РНК, выделенная из клеточных культур. Реакции ОТ-ПЦР и ПЦР в реальном времени проводили, как описано в разделе «Материалы и методы». ( A ) RT-PCR анализ E-NPPs, NTPDases, Ecto-5′-NT/CD73, ALP и ADA ( B ) Количественный анализ относительной экспрессии E-NPPs, NTPDases, Ecto-5′ -NT/CD73, ALP и ADA в клеточных линиях MB проводили с помощью ПЦР в реальном времени, где экспрессию GAPDH использовали в качестве внутреннего контроля для нормализации уровней экспрессии.Данные анализировали с помощью теста One-Way ANOVA с последующим апостериорным тестом Тьюки. Эксперименты проводили трижды в трехкратной повторности с (* p<0,01) и (*** p<0,001), что указывает на статистически значимое различие по сравнению с контрольными данными.
Реакции ОТ-ПЦР и ПЦР в реальном времени проводили, как описано в разделе «Материалы и методы». ( A ) RT-PCR анализ E-NPPs, NTPDases, Ecto-5′-NT/CD73, ALP и ADA ( B ) Количественный анализ относительной экспрессии E-NPPs, NTPDases, Ecto-5′ -NT/CD73, ALP и ADA в клеточных линиях MB проводили с помощью ПЦР в реальном времени, где экспрессию GAPDH использовали в качестве внутреннего контроля для нормализации уровней экспрессии.Данные анализировали с помощью теста One-Way ANOVA с последующим апостериорным тестом Тьюки. Эксперименты проводили трижды в трехкратной повторности с (* p<0,01) и (*** p<0,001), что указывает на статистически значимое различие по сравнению с контрольными данными.
https://doi.org/10.1371/journal.pone.0047468.g002
Статистический анализ
Данные выражали как среднее значение ± стандартное отклонение по крайней мере трех независимых экспериментов и подвергали однофакторному дисперсионному анализу (ANOVA) с последующим апостериорным тестом Тьюки или двусторонним дисперсионным анализом с последующим тестом Бонферрони или апостериорным тестом Тьюки, когда необходимый. Различия между средними значениями считались значимыми при p<0,05 .
Различия между средними значениями считались значимыми при p<0,05 .
Результаты
Секреция аденинового нуклеотида/нуклеозида клетками Daoy, ONS76 и D283 MB
клеток MB культивировали и использовали для определения секреции адениновых нуклеотидов/нуклеозидов. Все три проанализированные клеточные линии преимущественно секретируют АТФ во внеклеточную среду (рис. 1). Клеточная линия Daoy показала более высокую секрецию (100,04 мкмоль ± 14,99), чем клеточные линии ONS76 и D283 MB (55.61 мкмоль ±0,145 и 26,48 мкмоль ±10,54 соответственно). Важно отметить, что три клеточные линии МБ секретировали АДФ и АДО во внеклеточную среду, хотя и в гораздо более низких концентрациях, чем секретируемый АТФ. Различия в секреции между клеточными линиями MB не были статистически значимыми. Обнаруженная ИНО, вероятно, является результатом дезаминирования АДО присутствующей в этих клетках АДА.
Рисунок 3. Временная кинетика активности эктонуклеотидпирофосфатазы/фосфодиэстеразы в клеточных линиях МВ.
После достижения слияния клетки Daoy ( A ), ONS76 ( B ) и D283 ( C ) MB инкубировали с 0,5 мМ 5′-TMP-p-Nph, как описано в разделе «Материалы и методы». Значения представляют собой среднее значение ± стандартное отклонение из шести независимых экспериментов, проведенных в трех повторностях. Удельную активность выражали в нмоль ρ-нитрофенола/мин/мг белка.
https://doi.org/10.1371/journal.pone.0047468.g003
Оценка транскрипции мРНК эктонуклеотидазы
АТФ передает различные сигналы в соседние клетки, и эти эффекты контролируются активностью эктонуклеотидазы [15].Учитывая, что клеточные линии МБ секретируют АТФ во внеклеточную среду, транскрипция мРНК эктонуклеотидаз в этих клеточных линиях была подтверждена с помощью ОТ-ПЦР (рис. 2А) и ПЦР в реальном времени (рис. 2В). Мы заметили, что все клеточные линии MB экспрессировали только мРНК NTPDase5, но экспрессию генов NTPDase1, 2 и 3 обнаружить не удалось (рис. 2A). ПЦР-анализ в реальном времени показал выраженную экспрессию NTPDase5 в клетках D283 по сравнению с другими клеточными линиями (рис. 2В). Кроме того, в клеточной линии D283 обнаружены ампликоны E-NPP1, 2 и 3 ожидаемых размеров; ПЦР в реальном времени подтвердила, что эти профили экспрессии были значительно выше, чем в других клеточных линиях MB.Клеточные линии Daoy и ONS76 MB экспрессировали все три типа E-NPP, но в очень низких пропорциях, существенно не различаясь между собой.
2A). ПЦР-анализ в реальном времени показал выраженную экспрессию NTPDase5 в клетках D283 по сравнению с другими клеточными линиями (рис. 2В). Кроме того, в клеточной линии D283 обнаружены ампликоны E-NPP1, 2 и 3 ожидаемых размеров; ПЦР в реальном времени подтвердила, что эти профили экспрессии были значительно выше, чем в других клеточных линиях MB.Клеточные линии Daoy и ONS76 MB экспрессировали все три типа E-NPP, но в очень низких пропорциях, существенно не различаясь между собой.
Рис. 4. Гидролиз АТФ, АДФ и АМФ клеточными линиями MB человека.
Конфлюэнтные культуры клеток Daoy, ONS76 и D283 инкубировали с АТФ, АДФ или АМФ, как описано в разделе «Материалы и методы». Для клеток D283 использовали концентрацию 2,0 мМ и время инкубации 30 минут для всех субстратов, тогда как для клеточных линий Daoy и ONS76 использовали 1.Использовали 0 мМ и 30 минут инкубации с АТФ и АДФ и 2,0 мМ и 10 минут инкубации с АМФ. Удельную активность выражали в нмоль Pi/мин/мг белка. Столбцы представляют собой среднее значение ± стандартное отклонение четырех независимых экспериментов, проведенных в трех экземплярах. Данные сравнивали с помощью теста Two-Way ANOVA с последующим апостериорным тестом Бонферрони. (***) p<0,001 и (*) p<0,05 по сравнению с контрольными данными.
Данные сравнивали с помощью теста Two-Way ANOVA с последующим апостериорным тестом Бонферрони. (***) p<0,001 и (*) p<0,05 по сравнению с контрольными данными.
https://doi.org/10.1371/journal.pone.0047468.g004
Далее мы оценили транскрипцию мРНК ферментов, участвующих во внеклеточном метаболизме АТФ.Экспрессия мРНК экто-5’NT/CD73 в первичных клеточных линиях MB (Daoy и ONS76) была выше, чем во вторичной клеточной линии MB (D283). Клеточные линии Daoy и D283 экспрессировали мРНК ALP в аналогичных пропорциях, а клеточные линии ONS76 и D283 демонстрировали сходные профили экспрессии ADA, обе выше, чем у клеточной линии Daoy (рис. 2A и 2B).
Рисунок 5. Метаболизм внеклеточного AMP клеточными линиями MB.
Сливающиеся культуры клеток Daoy ( A ), ONS76 ( B ) и D283 ( C ) культивировали в 24-луночных планшетах до слияния и инкубировали со 100 мкМ AMP в 250 мкл инкубационной среды, как описано в разделе «Материалы». и методы.Аликвоты супернатанта собирали на 0, 5, 10, 20, 30, 60 и 90 мин и определяли уровни AMP, ADO, INO и HIPO (гипоксантина) с помощью ВЭЖХ. Данные выражали в виде средних значений ± стандартное отклонение из двух независимых экспериментов, проведенных в трех повторностях.
и методы.Аликвоты супернатанта собирали на 0, 5, 10, 20, 30, 60 и 90 мин и определяли уровни AMP, ADO, INO и HIPO (гипоксантина) с помощью ВЭЖХ. Данные выражали в виде средних значений ± стандартное отклонение из двух независимых экспериментов, проведенных в трех повторностях.
https://doi.org/10.1371/journal.pone.0047468.g005
Эктонуклеотидная активность пирофосфатазы/фосфодиэстеразы (E-NPP)
Принимая во внимание, что E-NPPs могут помочь поддерживать идеальные уровни нуклеотидов во внеклеточной среде [34] и что их экспрессия была обнаружена в проанализированных здесь клеточных линиях MB, мы оценили активность этих ферментов, как описано в разделе «Материалы и методы».Три изученные клеточные линии показали большую активность E-NPP при щелочном рН, оптимальный рН для этих ферментов in vitro [35]. Однако при физиологических значениях pH ферментативная активность E-NPP была чрезвычайно низкой в клеточных линиях MB, что указывает на его участие во внеклеточном метаболизме нуклеотидов даже в этих условиях (рис. 3).
3).
Рисунок 6. Специфичность субстрата.
После слияния клеточные линии Daoy, ONS76 и D283 MB инкубировали с различными монофосфонуклеозидами, как описано в разделе «Материалы и методы».Столбцы представляют собой среднее значение ± стандартное отклонение 6 независимых экспериментов, проведенных в трех экземплярах. Удельную активность выражали в нмоль Pi/мин/мг белка. Данные сравнивали с помощью теста Two-Way ANOVA с последующим апостериорным тестом Бонферрони. (***) p<0,001 и было принято для обозначения статистической значимости.
https://doi.org/10.1371/journal.pone.0047468.g006
Рисунок 7. Внеклеточный гидролиз фосфатных эфиров клеточными линиями MB.
После слияния клетки MB инкубировали и анализировали количество неорганического фосфата, как описано в разделе «Материалы и методы».( A ) β-глицерофосфат (β-Gli-P), глюкозо-6-фосфат (Gli-6-P) и неорганический пирофосфат (PPi), специфические субстраты для неспецифических фосфатаз, использовали для оценки ферментативной активности этих ферментов. . ( B ) Клетки МВ инкубировали с АМФ в присутствии левамизола, специфического ингибитора щелочной фосфатазы, для оценки влияния активности фосфатаз на гидролиз АМФ. Удельную активность выражали в нмоль Pi/мин/мг белка. Данные сравнивали с помощью теста Two-Way ANOVA с последующим апостериорным тестом Бонферрони.Все эксперименты проводились в трехкратной повторности.
. ( B ) Клетки МВ инкубировали с АМФ в присутствии левамизола, специфического ингибитора щелочной фосфатазы, для оценки влияния активности фосфатаз на гидролиз АМФ. Удельную активность выражали в нмоль Pi/мин/мг белка. Данные сравнивали с помощью теста Two-Way ANOVA с последующим апостериорным тестом Бонферрони.Все эксперименты проводились в трехкратной повторности.
https://doi.org/10.1371/journal.pone.0047468.g007
Гидролиз АТФ, АДФ и АМФ
Затем мы стремились проанализировать ферментативную активность E-NTPDase. С этой целью исследовали дифференциальную картину гидролиза АТФ, АДФ и АМФ в клеточных линиях Daoy, ONS76 и D283. Все клеточные линии МБ показали низкую скорость гидролиза АТФ и АДФ (рис. 4). Однако, когда мы проанализировали гидролиз AMP, ферментативная активность клеточных линий MB продемонстрировала специфический и дифференциальный профиль с Daoy > ONS76 > D283.Удельная активность составила 74,11 ± 8,33, 9,206 ± 0,730 и 3,654 ± 0,157 нмоль Pi/мин/мг белка, соответственно, демонстрируя самую высокую активность AMPase в клеточной линии Daoy.
Рисунок 8. Анализ экспрессии белка экто-5′-NT/CD73 с помощью проточной цитометрии в клеточных линиях Daoy, ONS76 и D283 MB.
Конфлюэнтные клеточные линии МВ обрабатывали трипсином и готовили, как описано в разделе «Материалы и методы». После инкубации с очищенным мышиным антителом против человеческого CD73 (1∶10) и мечением Alexa fluor 555 Rabbit против мышиных IgG (1∶100) клетки анализировали с помощью проточной цитометрии.( A ) Результаты представлены в виде соотношения меченых и немеченых клеток. ( B ) Репрезентативное изображение экто-5’NT/CD73-позитивных клеток. Статистическую релевантность анализировали с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Тьюки. Эксперименты проводились в трехкратной повторности. (**) p<0,001 по сравнению с контролем.
https://doi.org/10.1371/journal.pone.0047468.g008
Анализ внеклеточного метаболизма AMP методом ВЭЖХ
Активность АМФазы клеточных линий МБ дополнительно изучали с помощью ВЭЖХ. На фигуре 5 показано, что AMP эффективно гидролизовался клеточными линиями Daoy и ONS76 MB. Эти клетки почти полностью гидролизуют АМП через 30 мин инкубации; однако профили гидролиза были разными. В то время как Daoy превращал AMP в ADO и накапливал эту молекулу в конце времени инкубации, ONS76 продуцировал ADO, который, вероятно, превращался в INO; эта молекула накапливалась как основной продукт метаболизма АМФ. Подтверждая результаты, полученные с помощью колориметрического анализа (рис. 4), клеточная линия D283 MB не гидролизовала AMP.
На фигуре 5 показано, что AMP эффективно гидролизовался клеточными линиями Daoy и ONS76 MB. Эти клетки почти полностью гидролизуют АМП через 30 мин инкубации; однако профили гидролиза были разными. В то время как Daoy превращал AMP в ADO и накапливал эту молекулу в конце времени инкубации, ONS76 продуцировал ADO, который, вероятно, превращался в INO; эта молекула накапливалась как основной продукт метаболизма АМФ. Подтверждая результаты, полученные с помощью колориметрического анализа (рис. 4), клеточная линия D283 MB не гидролизовала AMP.
Рисунок 9. Цитозольная и ядерная экспрессия P-β-катенина в клеточных линиях MB.
Экспрессию фосфо-β-катенина определяли с помощью ( A ) иммунофлуоресцентного анализа и ( B ) вестерн-блоттинга, как описано в разделе «Материалы и методы». Для иммунофлуоресценции изображения получали с помощью объектива с 20-кратным увеличением, а цитозольную и ядерную иммунореактивность фосфо-β-катенина наблюдали в колонке слияния. Для вестерн-блоттинга относительные уровни фосфо-β-катенина цитозольной и ядерной фракций, полученные с помощью денсиметрического анализа белковых полос, обнаруженных в вестерн-блоттинге, сравнивали с уровнями экспрессии GAPDH и HnRNPK соответственно.Столбцы представляют собой среднее значение ± стандартное отклонение 3 независимых экспериментов, проведенных в трех экземплярах. Данные сравнивали с помощью теста One-Way ANOVA с последующим апостериорным тестом Тьюки. (*р<0,01) по сравнению с контрольными данными.
Для вестерн-блоттинга относительные уровни фосфо-β-катенина цитозольной и ядерной фракций, полученные с помощью денсиметрического анализа белковых полос, обнаруженных в вестерн-блоттинге, сравнивали с уровнями экспрессии GAPDH и HnRNPK соответственно.Столбцы представляют собой среднее значение ± стандартное отклонение 3 независимых экспериментов, проведенных в трех экземплярах. Данные сравнивали с помощью теста One-Way ANOVA с последующим апостериорным тестом Тьюки. (*р<0,01) по сравнению с контрольными данными.
https://doi.org/10.1371/journal.pone.0047468.g009
Гидролиз других монофосфонуклеозидов
Подобно гидролизу АМФ, клеточные линии MB продемонстрировали ту же картину гидролиза других монофосфонуклеозидов (CMP, GMP, IMP и UMP) (рис. 6), как и ожидалось, для экто-5’NT/CD73 [17].Как было показано ранее, Daoy и D283 экспрессируют мРНК ALP, еще одного фермента, который может гидролизовать AMP и другие эфиры фосфорной кислоты. Daoy и ONS76, которые демонстрировали более высокую активность AMPase, показали очень низкую ферментативную активность в присутствии Glu-6P или β-Gly-P, двух субстратов для ALP (рис. 7A). Клетки D283 MB также плохо гидролизовали эти субстраты. Кроме того, потенциальное участие активности ALP в гидролизе AMP оценивали по присутствию левамизола, специфического ингибитора этого фермента (фиг. 7B).Активность АМФ в клеточных линиях МБ не изменялась в присутствии ингибитора, за исключением участия ЩФ в гидролизе АМФ. Мы наблюдали заметный гидролиз PPi всеми клеточными линиями МВ (рис. 7А). Активность неорганической пирофосфатазы также была обнаружена в трех клеточных линиях (рис. 7А), что согласуется с присутствием экто-пирофосфатазы во всех проанализированных здесь клеточных линиях MB.
7A). Клетки D283 MB также плохо гидролизовали эти субстраты. Кроме того, потенциальное участие активности ALP в гидролизе AMP оценивали по присутствию левамизола, специфического ингибитора этого фермента (фиг. 7B).Активность АМФ в клеточных линиях МБ не изменялась в присутствии ингибитора, за исключением участия ЩФ в гидролизе АМФ. Мы наблюдали заметный гидролиз PPi всеми клеточными линиями МВ (рис. 7А). Активность неорганической пирофосфатазы также была обнаружена в трех клеточных линиях (рис. 7А), что согласуется с присутствием экто-пирофосфатазы во всех проанализированных здесь клеточных линиях MB.
Анализ экспрессии белка экто-5’NT/CD73 методом проточной цитометрии
Чтобы подтвердить присутствие экто-5’NT/CD73 на клеточной поверхности, мы провели анализ методом проточной цитометрии, как описано в разделе «Материалы и методы».Фигура 8А подтверждает данные, представленные в анализе экспрессии мРНК, где клеточные линии, представляющие первичный MB (Daoy и ONS76), показали более высокую экспрессию экто-5’NT/CD73, чем клеточная линия MB D283, представляющая вторичный MB.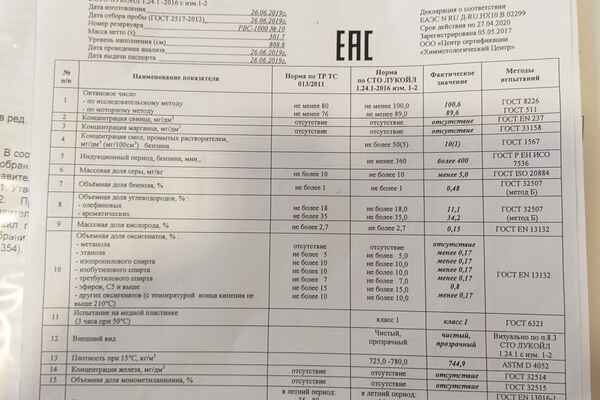 Результаты были дополнительно оценены количественно и представлены следующими процентами экспрессии мембраны экто-5’NT/CD73 в каждой клеточной линии MB: Daoy: 74,52% ±8,026, ONS76∶67,97±4,087 и D283∶3,115±1,393.
Результаты были дополнительно оценены количественно и представлены следующими процентами экспрессии мембраны экто-5’NT/CD73 в каждой клеточной линии MB: Daoy: 74,52% ±8,026, ONS76∶67,97±4,087 и D283∶3,115±1,393.
Экспрессия клеток фосфо-β-катенина МВ
β-катенин является нижестоящим эффектором канонического пути Wnt и, как описано ранее, важен для прогноза и определения стадии опухоли MB [3], [5].Мы использовали анализы иммунофлуоресценции и вестерн-блоттинга для обнаружения экспрессии фосфо-β-катенина в используемых здесь клеточных линиях MB. Во-первых, мы заметили, что Daoy и ONS76 проявляли в основном ядерную иммунореактивность к фосфо-β-катенину, в то время как клеточная линия D283 MB проявляла иммунореактивность к этому белку в цитозоле (рис. 9А). Эти качественные данные были подтверждены вестерн-блоттингом цитозольной и ядерной фракций по сравнению с их соответствующими контролями, GAPDH и HnRNPK (фиг. 9B).
Обсуждение
В настоящей работе мы изучили экспрессию и активность эктонуклеотидаз в трех различных клеточных линиях медуллобластомы (МБ) (Daoy, ONS76 и D283). АТФ высвобождается во внеклеточную среду при физиологических или патологических состояниях, стимулируя различные типы сигнальных путей [12]. Анализируемые клеточные линии МБ секретировали АТФ во внеклеточную среду в микромолярном количестве. Полученные данные согласуются с предыдущими результатами нашей группы с клеточной линией глиомы U138MG, которая также секретировала АТФ [36]. Кроме того, внеклеточный АТФ в концентрации 100 мкМ стимулировал пролиферацию клеток глиомы [26], будучи цитотоксичным для культур срезов органотипического гиппокампа крысы [24].
АТФ высвобождается во внеклеточную среду при физиологических или патологических состояниях, стимулируя различные типы сигнальных путей [12]. Анализируемые клеточные линии МБ секретировали АТФ во внеклеточную среду в микромолярном количестве. Полученные данные согласуются с предыдущими результатами нашей группы с клеточной линией глиомы U138MG, которая также секретировала АТФ [36]. Кроме того, внеклеточный АТФ в концентрации 100 мкМ стимулировал пролиферацию клеток глиомы [26], будучи цитотоксичным для культур срезов органотипического гиппокампа крысы [24].
Попадая во внеклеточную среду, АТФ подвергается деградации эктонуклеотидазами. Представленные здесь результаты показывают, что клеточные линии MB не экспрессируют E-NTPDases, за исключением NTPDase5. Эти данные согласуются с данными о злокачественных глиомах, которые демонстрируют очень низкую активность АТФазы и АДФазы по сравнению с их нормальными аналогами, астроцитами [23]. Наличие доминирующей NTPDase2 в первичных культурах астроцитов гиппокампа, коры и мозжечка [25] может указывать на потерю экспрессии/активности E-NTPDase при злокачественной трансформации опухолей головного мозга. Точно так же NTPDase5, внутриклеточно экспрессируемый фермент, который считается протоонкогеном, отсутствует в нормальных астроцитах. Следовательно, скорость его экспрессии может коррелировать со степенью злокачественности [11], [37].
Точно так же NTPDase5, внутриклеточно экспрессируемый фермент, который считается протоонкогеном, отсутствует в нормальных астроцитах. Следовательно, скорость его экспрессии может коррелировать со степенью злокачественности [11], [37].
E-NPP также гидролизуют ди- и трифосфатные нуклеотиды во внеклеточной среде. Различные типы тканей экспрессируют эти ферменты, которые связаны с различными патофизиологическими и биохимическими функциями [35], [38]. Здесь мы демонстрируем, что Daoy и ONS76 обнаруживают низкую экспрессию и активность всех проанализированных E-NPP по сравнению с клеточной линией D283.Клетки D283 экспрессировали все E-NPP в следующем порядке: NPP2> NPP1> NPP3. Экспрессия NPP3 наблюдалась в незрелых астроцитах пренатальных и неонатальных крыс, взаимодействуя с другими белками для трансформации опухоли глиальных клеток после обработки N,N’-этилнитрозомочевиной [35]. Следовательно, наша гипотеза состоит в том, что экспрессия этого фермента коррелирует со стадией недифференцированной клетки. NPP1, предполагаемый маркер злокачественности глиомы, присутствовал в клеточной линии крысиной глиомы C6, где вместе с NPP3 отвечал за гидролиз низких концентраций АТФ в клеточной линии глиомы C6 (1–10 мкМ) [39].В исследовании мы показываем, что активность E-NPP также присутствовала на низких уровнях, судя по скорости гидролиза его специфического искусственного субстрата, 5′-TMP-p-Nph. Эти результаты вместе с отсутствием экспрессии E-NTPDase согласуются с низкой скоростью гидролиза нуклеотидов АТФ и АДФ, и это может быть связано с активностью E-NPP.
NPP1, предполагаемый маркер злокачественности глиомы, присутствовал в клеточной линии крысиной глиомы C6, где вместе с NPP3 отвечал за гидролиз низких концентраций АТФ в клеточной линии глиомы C6 (1–10 мкМ) [39].В исследовании мы показываем, что активность E-NPP также присутствовала на низких уровнях, судя по скорости гидролиза его специфического искусственного субстрата, 5′-TMP-p-Nph. Эти результаты вместе с отсутствием экспрессии E-NTPDase согласуются с низкой скоростью гидролиза нуклеотидов АТФ и АДФ, и это может быть связано с активностью E-NPP.
NPP2 выполняет многочисленные функции, включая стимулирование ангиогенеза, пролиферации и дифференцировки клеток. NPP2 был наиболее известным ферментом E-NPP, экспрессируемым клеточной линией D283, полученной из метастатической опухоли.Это наблюдение согласуется с ранее опубликованными сообщениями, предполагающими функции NPP2 как стимулятора подвижности опухоли в клетках меланомы и при раке молочной железы. Кроме того, экспрессия этого фермента повышалась при раке легкого и зависела от степени роста и инвазивности [35], [40]. Эти данные, связанные с секрецией АТФ, представленной клеточными линиями MB, согласуются с современной литературой, предполагающей, что АТФ накапливается во внеклеточном пространстве клеток MB.
Эти данные, связанные с секрецией АТФ, представленной клеточными линиями MB, согласуются с современной литературой, предполагающей, что АТФ накапливается во внеклеточном пространстве клеток MB.
Важным открытием нашей настоящей работы является то, что первичные опухолевые клетки MB, такие как линии Daoy и ONS76, экспрессируют экто-5’NT/CD73, в то время как клеточная линия D283, представляющая метастатическую опухоль, показала очень низкую экспрессию этого фермента.Эти результаты интригуют, учитывая, что повышенная экспрессия/активность экто-5’NT/CD73 является наиболее очевидным изменением эктонуклеотидазного пути, связанным со степенью злокачественности различных типов опухолей, включая глиомы [8]. Кроме того, мы показали, что три клеточные линии MB экспрессируют экто-аденозиндезаминазу (ADA) в порядке, обратном профилю экспрессии экто-5’NT/CD73, что подтверждается различиями в продукции аденозина и инозина, как показано в Рисунок 5.
Возможное объяснение этой инверсии паттернов экспрессии/активности экто-5’NT/CD73 и ADA в клетках МВ может быть обеспечено путем пути Wnt/β-катенин. Передача сигнала, индуцированная Wnt/β-catenin, регулирует экспрессию нескольких генов, связанных с раком, включая MB [41]. Вкратце, цитоплазматические уровни β-катенина могут контролироваться путем Wnt, поскольку он может дестабилизировать мультимерный белковый комплекс в цитоплазме, приводя к повышенным уровням свободного β-катенина, который подвергается ядерной транслокации. В неактивном состоянии пути GSK-3β и APC, оба присутствующие в цитозольном белковом комплексе, фосфорилируют цитозольный β-катенин, который затем становится убиквитинированным для последующей цитоплазматической деградации [7], [42].Мутации APC были обнаружены при самых разных раковых заболеваниях человека, включая MB. Недостаточное фосфорилирование β-катенина с помощью APC нарушает его цитоплазматическую деградацию и, следовательно, способствует транслокации фосфо-β-катенина в ядро [42]. Кроме того, цитоплазматическая иммунореактивность на β-катенин в образцах опухолевой ткани пациентов с МБ была связана с плохим прогнозом, тогда как ядерная иммунореактивность на β-катенин была связана с лучшим прогнозом, при этом пациенты демонстрировали большую продолжительность жизни [7].
Передача сигнала, индуцированная Wnt/β-catenin, регулирует экспрессию нескольких генов, связанных с раком, включая MB [41]. Вкратце, цитоплазматические уровни β-катенина могут контролироваться путем Wnt, поскольку он может дестабилизировать мультимерный белковый комплекс в цитоплазме, приводя к повышенным уровням свободного β-катенина, который подвергается ядерной транслокации. В неактивном состоянии пути GSK-3β и APC, оба присутствующие в цитозольном белковом комплексе, фосфорилируют цитозольный β-катенин, который затем становится убиквитинированным для последующей цитоплазматической деградации [7], [42].Мутации APC были обнаружены при самых разных раковых заболеваниях человека, включая MB. Недостаточное фосфорилирование β-катенина с помощью APC нарушает его цитоплазматическую деградацию и, следовательно, способствует транслокации фосфо-β-катенина в ядро [42]. Кроме того, цитоплазматическая иммунореактивность на β-катенин в образцах опухолевой ткани пациентов с МБ была связана с плохим прогнозом, тогда как ядерная иммунореактивность на β-катенин была связана с лучшим прогнозом, при этом пациенты демонстрировали большую продолжительность жизни [7].
Экспрессия экто-5’NT/CD73 и ADA обратно регулируется взаимодействиями между β-катенином и TCF/LEF, в результате чего образуется комплекс ядерных белков, который взаимодействует с промоторной областью этих генов. Соответственно, трансфекция генов β-катенина и TCF-1 в клетках Rat-1 усиливала экто-5’NT/CD73 и снижала активность и экспрессию ADA [43]. Наши результаты показали, что ядерная иммунореактивность β-катенина была выше в клеточных линиях Daoy и ONS76, чем в клетках D283. Учитывая, что метастатический фенотип связан с плохим прогнозом для пациента [41], мы предполагаем, что отсутствие экто-5’NT/CD73 и присутствие ADA в клеточной линии D283, метастатическом фенотипе MB, могут быть связаны с его плохим прогноз.Необходимы дальнейшие исследования, чтобы полностью выяснить участие этих ферментов в прогрессировании MB.
Благодарности
Мы благодарим Д. Ямамото и Л.Р. Blazina за их техническую помощь; д-р C. Lameu за помощь в анализе количественной ПЦР; R. Dariolli для фосфо-β-катенина и Drs. G. Schwartsmann и A. Brunetto для клеточных линий медуллобластомы.
Dariolli для фосфо-β-катенина и Drs. G. Schwartsmann и A. Brunetto для клеточных линий медуллобластомы.
Авторские взносы
Задумал и спроектировал эксперименты: ARC LR FD VC TG EB ALA RR HU AMOB.Проведены эксперименты: АРК ЛР ФД ВК ТГ. Проанализированы данные: ARC. Предоставленные реагенты/материалы/инструменты для анализа: ARC LR FD VC TG EB ALA RR HU AMOB. Написал статью: ARC AMOB.
Каталожные номера
- 1. Koeller KK, Rushing EJ (2003) Из архивов AFIP: медуллобластома: всесторонний обзор с радиологической и патологической корреляцией. Рентгенография 23: 1613–1637.
- 2. Muoio VM, Shinjo SO, Matushita H, Rosemberg S, Teixeira MJ, et al.(2011)Экстранейральные метастазы медуллобластомы. Arq Neuropsiquiatr 69: 328–331.
- 3. Луи Д.Н., Огаки Х., Вистлер О.Д., Кавени В.К., Бургер П.С. и др. (2007) Классификация ВОЗ опухолей центральной нервной системы 2007 года. Acta Neuropathol 114: 97–109.
- 4. Schmidt AL, Brunetto AL, Schwartsmann G, Roesler R, Abujamra AL (2010)Последние терапевтические достижения в лечении медуллобластомы: фокус на новых молекулярных мишенях. Лекарственные препараты для лечения нервных расстройств ЦНС 9: 335–348.
- 5. Руссель М.Ф., Робинсон Г. (2011)Медуллобластома: достижения и проблемы. F1000 Биол Реп 3: 5.
- 6. Rodini CO, Suzuki DE, Nakahata AM, Pereira MC, Janjoppi L, et al. (2010)Аберрантные сигнальные пути в медуллобластомах: связь со стволовыми клетками. Arq Neuropsiquiatr 68: 947–952.
- 7. Эллисон Д.В., Онилюд О.Е., Линдси Дж.К., Лушер М.Е., Уэстон С.Л. и др. (2005) Статус бета-катенина предсказывает благоприятный исход медуллобластомы у детей: Комитет по опухолям головного мозга Детской исследовательской группы рака Соединенного Королевства.Дж. Клин Онкол 23: 7951–7957.
- 8.
Spychala J (2000)Опухолестимулирующие функции аденозина. Pharmacol Ther 87: 161–173.
- 9. Бавареско Л., Бернарди А., Браганхол Э., Капеллари А.Р., Рокенбах Л. и др. (2008)Роль экто-5′-нуклеотидазы/CD73 в пролиферации клеточной линии глиомы. Mol Cell Biochem 319: 61–68.
- 10. Stagg J, Smyth MJ (2010)Внеклеточный аденозинтрифосфат и аденозин при раке. Онкоген 29: 5346–5358.
- 11.Стелла Дж., Бавареско Л., Браганхол Э., Рокенбах Л., Фариас П.Ф. и др. (2009)Дифференциальная экспрессия эктонуклеотидазы в клеточных линиях рака мочевого пузыря человека. Урол Онкол 28: 260–267.
- 12. Филдс Р.Д., Бернсток Г. (2006)Пуринергическая передача сигналов во взаимодействиях нейронов и глии. Nat Rev Neurosci 7: 423–436.
- 13. Abbrachio MP, Burnstock G, Verkhratsky A, Zimmermann H (2009)Пуринергическая передача сигналов в нервной системе: обзор. Тенденции Neurosci 32: 19–29.
- 14.
Robson SC, Sevigny J, Zimmermann H (2006)Семейство E-NTPDase эктонуклеотидаз: структурно-функциональные отношения и патофизиологическое значение. Пуринергический сигнал 2: 409–430.
- 15. Zimmermann H (2000)Внеклеточный метаболизм АТФ и других нуклеотидов. Naunyn Schmiedebergs Arch Pharmacol 362: 299–309.
- 16. Циммерманн Х., Зебиш М., Стратер Н. (2012)Клеточная функция и молекулярная структура эктонуклеотидаз.Пуринергический сигнал 8: 437–502.
- 17. Strater N (2006)Экто-5′-нуклеотидаза: взаимоотношения структурных функций. Пуринергический сигнал 2: 343–350.
- 18. Егуткин Г.Г. (2008) Нуклеотид- и нуклеозидпревращающие эктоферменты: важные модуляторы пуринергического сигнального каскада. Биохим Биофиз Акта 1783: 673–694.
- 19. Millan JL (2006) Щелочные фосфатазы: структура, субстратная специфичность и функциональное родство с другими членами большого надсемейства ферментов.Пуринергический сигнал 2: 335–341.
- 20.
Окубо С., Кимура Дж., Мацуока И. (2000)Экто-щелочная фосфатаза в клетках NG108–15: ключевой фермент, опосредующий чувствительный к антагонисту P1 ответ АТФ. Br J Pharmacol 131: 1667–1672.
- 21. Picher M, Burch LH, Hirsh AJ, Spychala J, Boucher RC (2003)Экто-5′-нуклеотидаза и неспецифическая щелочная фосфатаза. Два AMP-гидролизующих эктофермента с разными ролями в дыхательных путях человека. J Biol Chem 278: 13468–13479.
- 22.Франко Р., Касадо В., Сируэла Ф., Саура С., Маллол Дж. и др. (1997) Аденозиндезаминаза клеточной поверхности: гораздо больше, чем эктофермент. Прог Нейробиол 52: 283–294.
- 23. Wink MR, Lenz G, Braganhol E, Tamajusuku AS, Schwartsmann G, et al. (2003)Измененный внеклеточный катаболизм АТФ, АДФ и АМФ в клеточных линиях глиомы. Рак Летт 198: 211–218.
- 24. Морроне Ф.Б., Хорн А.П., Стелла Дж., Спиллер Ф., Саркис Дж.Дж. и др. (2005)Повышенная устойчивость клеточных линий глиомы к внеклеточной цитотоксичности АТФ.Дж. Нейронкол 71: 135–140.
- 25.
Wink MR, Braganhol E, Tamajusuku AS, Casali EA, Karl J, et al. (2003) Метаболизм внеклеточных адениновых нуклеотидов в культурах астроцитов из разных областей мозга. Нейрохим Инт 43: 621–628.
- 26. Морроне Ф.Б., Жак-Сильва М.С., Хорн А.П., Бернарди А., Швартсманн Г. и др. (2003) Внеклеточные нуклеотиды и нуклеозиды индуцируют пролиферацию и увеличивают транспорт нуклеозидов в клеточных линиях глиомы человека. Дж. Нейронкол 64: 211–218.
- 27. Кастеллино Р.С., Де Бортоли М., Лин Л.Л., Скапура Д.Г., Раджан Дж.А. и др. (2007)Сверхэкспрессия TP73 индуцирует апоптоз в медуллобластоме. BMC Рак 7: 127.
- 28. Хау А.К., Силасуджана М., Банерджи Б., Судзуки М., Баскар Р. и др. (2007)Ингибирование активности теломеразы и экспрессии гена обратной транскриптазы теломеразы человека ингибитором гистондеацетилазы в клетках рака головного мозга человека. Мутат Рез 625: 134–144.
- 29. Belcher SM, Ma X, Le HH (2009)Блокада передачи сигналов рецептора эстрогена подавляет рост и миграцию медуллобластомы.Эндокринология 150: 1112–1121.
- 30.
фон Бюрен А.О., Шалаби Т., Олер-Янне С. , Арнольд Л., Стернс Д. и др. (2009) Ингибирование c-MYC, опосредованное РНК-интерференцией, предотвращает рост клеток и снижает чувствительность к радио- и химиотерапии в клетках медуллобластомы у детей. BMC Рак 9: 10.
- 31. Sakura H, Nagashima S, Nakashima A, Maeda M (1998) Характеристика 5′-нуклеотидной фосфодиэстеразы сыворотки плода: новая функция ингибитора агрегации тромбоцитов в кровообращении плода.Тромб Рез. 91: 83–89.
- 32. Чан К.М., Дельферт Д., Юнгер К.Д. (1986) Прямой колориметрический анализ Са2+-стимулируемой АТФазной активности. Анальный биохим 157: 375–380.
- 33. Bradford MM (1976) Быстрый и чувствительный метод количественного определения количества белка в микрограммах, использующий принцип связывания белка с красителем. Анальный биохим 72: 248–254.
- 34.
Стефан С., Янсен С., Боллен М. (2006) Модуляция пуринергической передачи сигналов эктофосфодиэстеразами типа NPP.Пуринергический сигнал 2: 361–370.
- 35. Goding JW, Grobben B, Slegers H (2003)Физиологические и патофизиологические функции семейства эктонуклеотидных пирофосфатаз/фосфодиэстераз. Биохим Биофиз Акта 1638: 1–19.
- 36. Braganhol E, Tamajusuku AS, Bernardi A, Wink MR, Battastini AM (2007)Ингибирование экто-5′-нуклеотидазы/CD73 кверцетином в клеточной линии глиомы человека U138MG. Биохим Биофиз Акта 1770: 1352–1359.
- 37. Вильяр Дж., Аренас М.И., Маккарти К.М., Бланкес М.Дж., Тирадо О.М. и др.(2007) Экспрессия PCPH/ENTPD5 усиливает инвазивность клеток рака предстательной железы человека с помощью механизма, зависящего от протеинкиназы C-дельта. Рак Res 67: 10859–10868.
- 38. Aerts I, Martin JJ, De Deyn PP, Van Ginniken C, Van Ostade X и др. (2011) Экспрессия эктонуклеотидной пирофосфатазы/фосфодиэстеразы 1 (E-NPP1) коррелирует со степенью астроцитарной опухоли. Клин Нейрол Нейросург 113: 224–229.
- 39.
Гроббен Б., Ансьо К. , Ройманс Д., Стефан С., Боллен М. и др.(1999) Эктонуклеотидная пирофосфатаза является одним из основных ферментов, участвующих во внеклеточном метаболизме АТФ в крысиной глиоме С6. Дж. Нейрохим 72: 826–834.
- 40. Стефан С., Янсен С., Боллен М. (2005)Эктофосфодиэстеразы типа NPP: единство в разнообразии. Trends Biochem Sci 30: 542–550.
- 41. Gilbertson RJ, Ellison DW (2008)Происхождение подтипов медуллобластомы. Анну Рев Патол 3: 341–365.
- 42. Polakis P (2000)Передача сигналов Wnt и рак.Гены Дев 14: 1837–1851.
- 43. Spychala J, Kitajewski J (2004) Wnt и передача сигналов бета-катенина нацелены на экспрессию экто-5′-нуклеотидазы и увеличивают внеклеточную генерацию аденозина. Разрешение ячейки опыта 296: 99–108.
 Schmidt AL, Brunetto AL, Schwartsmann G, Roesler R, Abujamra AL (2010)Последние терапевтические достижения в лечении медуллобластомы: фокус на новых молекулярных мишенях. Лекарственные препараты для лечения нервных расстройств ЦНС 9: 335–348.
Schmidt AL, Brunetto AL, Schwartsmann G, Roesler R, Abujamra AL (2010)Последние терапевтические достижения в лечении медуллобластомы: фокус на новых молекулярных мишенях. Лекарственные препараты для лечения нервных расстройств ЦНС 9: 335–348. Br J Pharmacol 131: 1667–1672.
Br J Pharmacol 131: 1667–1672. Нейрохим Инт 43: 621–628.
Нейрохим Инт 43: 621–628. , Арнольд Л., Стернс Д. и др. (2009) Ингибирование c-MYC, опосредованное РНК-интерференцией, предотвращает рост клеток и снижает чувствительность к радио- и химиотерапии в клетках медуллобластомы у детей. BMC Рак 9: 10.
, Арнольд Л., Стернс Д. и др. (2009) Ингибирование c-MYC, опосредованное РНК-интерференцией, предотвращает рост клеток и снижает чувствительность к радио- и химиотерапии в клетках медуллобластомы у детей. BMC Рак 9: 10.Обновление Hex.pm до Ecto 3.0 « Блог Plataformatec
Ecto 3.0 не за горами, и, как вы, возможно, уже знаете, он достиг стабильного API. Чтобы убедиться, что все работает правильно, я решил попробовать обновить один из первых проектов, которые когда-либо использовали Ecto: Hex. вечера.
Все обновление было выполнено в одном запросе на вытягивание, который мы рассмотрим ниже.
Во-первых, необходимые шаги:
- Обновите mix.exs, чтобы он зависел от
ecto_sql, и увеличьте зависимость postgrex. Примечание. Обработка SQL была выделена в отдельный проектecto_sql, поэтому нам нужно добавить эту новую зависимость. (6b3b78cf) - DBConnection 2.0 больше не поставляется с пулами Sojourn и Poolboy, поэтому мы можем удалить конфигурацию пула
760026f3) - Говоря о пулах, мы должны убедиться, что
pool_sizeне менее2при выполнении миграции. Библиотека - JSON теперь установлена на адаптере, а не на Ecto (
e16ebd8f), и поскольку мы уже использовали рекомендуемый пакет, Джейсон, нам больше не нужна эта конфигурация. (66f9cbdf) - Ecto 3. 0 делает обработку даты/времени более строгой в отношении точности. Поэтому нам нужно либо обновить типы полей нашей схемы, либо убедиться, что мы обрезаем значения даты/времени.Например, когда мы определяем поле как
время, мы не можем поместить значение с микросекундной точностью, и точно так же мы не можем поместить в полеtime_usecзначение без микросекундной точности. (2e34b833) - Функции обработки ограничений, такие как
Ecto.Changeset.unique_constraint/3, теперь включают в метаданные ошибки тип и имя ограничения, что нарушило наш слишком конкретный тест. (3d19f903)
Во-вторых, мы получили пару предупреждений об устаревании, так что вот исправления:
- Теперь адаптер устанавливается при определении модуля репо, а не в конфигурации.(
д3911953) -
Ecto.Multi.run/3теперь принимает функцию 2-арности (первым аргументом теперь является репо) вместо функции 1-арности раньше. ( 95d11cc2)
 (
( Наконец, было несколько незначительных глюков (или дублирований!) специфичных для Hex.pm: c4168977 , 21eb0bf8 и 0929cd9e .
В целом процесс обновления был довольно простым. По пути было несколько мелких ошибок, которые были оперативно исправлены выше по течению.Ранее обновив Hex.pm до Ecto 2.0, что заняло несколько месяцев (мы начали его на раннем этапе, что сделало его быстро меняющейся целью), я действительно могу оценить уровень зрелости, которого достиг Ecto, и то, как легко было обновить на этот раз. 🙂
Обновление: добавлено примечание о pool_size при выполнении миграции.
Мы рады быть активными участниками процесса, внося свой вклад в экосистему и помогая компаниям внедрять Elixir на разных этапах их цикла внедрения.Мы тоже хотели бы работать с вами!
Считаете ли вы, что ваша компания могла бы извлечь выгоду из нашего опыта и помощи? Просто нажмите кнопку ниже..jpg)
Теги: ecto, elixir, hex, Опубликовано на английском языке, Комментарии к обновлению Hex.pm до Ecto 3.0
отключены
Посетите нас у новоотвореной бензиновой поставкой на Реметинечка цеста брой 151 край НОВА тв, где Вас чека любит и профессионально особь ,,,а у продажи од куповине освьежите себе у новоотвореном ресторане Белла у склопу укус.
Драги купчи,
jer ste Vi to tražili…
ЭКТО НЕДЕЛЬЯ НА ЛУКОЙЛ-У!!!
Nedeljom pa nadalje, svaku nedjelju na LUKOIL benzinskim postajama označenim ECTO znakom, vrhunska ECTO goriva po cijeni standardnih goriva.
Ваш ЛУКОЙЛ
На ЛУКОЙЛ бензинским постама у Республиканской Хорватской Вриеде найстрожи стандарти квалитете горива.
ЛУКОЙЛ ямчи врхунску квалитету своих производств ЭКТО Дизель, ЭКТО ПЛЮС, Евродизель БС, Евросупер 95 БС, УНП автопл.
Посебна нам е часть представити Вам ЭКТО бренда горит из ЛУКОЙЛ-а (ЭКТО ПЛЮС — евросупер и ЭКТО Дизель — евродизель БС), инновативна горит, произведена у склада по наиболее европейским стандартам. Kvaliteta ECTO goriva potvrđena je od strane stručnjaka, kao i od vas, naših vjernih kupaca.
ЭКТО ДИЗЕЛЬ ЕВРОДИЗЕЛЬ BS — НАВЫКНИ СЕ БИТИ ЯЧИ!
Желите свой автомобиль учинити снажний?
Напитайте газ новым горивом ECTO Diesel Eurodiesel BS.
ECTO Diesel Eurodiesel BS osigurava:
- povećanje snage motora do 9%;
- уштеда горива до 5%;
- побольшанье хладног старта;
- smanjenje buke i vibracija.
ЭКТО ПЛЮС ЕВРОСУПЕР 95 БС — НЕОБУЗДАНА СТРАСТ!
Маштате или брзини?
Напитайте свой автомобиль горивым ECTO PLUS EUROSUPER 95 BS и питайте его узкую динамику.
- Odlična kvaliteta goriva.
- Povećanje snage motora.
- Povećanje učinkovitosti rada motora.
- Штите мотор от сваранья насоса и корозие горивног сустава.
- Smanjenje potrošnje goriva.
Дефицит CD73/экто-5′-нуклеотидазы у мышей усиливает острую реакцию «трансплантат против хозяина» | Кровь
Пациенты с гематологическими злокачественными новообразованиями, рефрактерными к традиционной химиотерапии, имеют шанс на излечение с помощью аллогенной трансплантации гемопоэтических стволовых клеток (алло-HCT). 1 Однако успех этого лечения ограничен РТПХ. 2 Недавно мы показали, что внеклеточный АТФ, который высвобождается из умирающих или подвергшихся стрессу клеток и служит эндогенным сигналом опасности, вызывающим системные воспалительные реакции, усиливает РТПХ за счет активации пуринергического рецептора P2X 7 R. 3,4 Количество внеклеточного АТФ регулируется эктонуклеотидазами, такими как CD39, которая дефосфорилирует АТФ до АДФ и АМФ. Внеклеточный AMP дефосфорилируется до аденозина под действием CD73, гликозилфосфатидилинозитол-заякоренного гликопротеина с ферментативной активностью экто-5′-нуклеотидазы. 5-7 CD73 экспрессируется на многих типах клеток, включая субпопуляции Т-лимфоцитов, эндотелиальных и эпителиальных клеток. 7,8 Он также присутствует в виде секретируемой формы, в которой отсутствует гликозилфосфатидилинозитоловый якорь. 9 Аденозин, генерируемый CD73, может активировать любой из 4 связанных с G-белком 7-трансмембранных трансмембранных аденозиновых рецепторов (ARs; A 1 , A 2A , A 2B и A 3 90 ) и может действовать в качестве про- или противовоспалительного медиатора в зависимости от физиологических условий и типа задействованного АР. 10-12 В большинстве случаев активация рецепторов A 1 и A 3 является провоспалительной, тогда как активация рецепторов A 2A и A 2B является противовоспалительной или толерогенной. 13,14
Важность CD73 в производстве аденозина для передачи сигналов AR была выявлена в ходе исследований на мышах с дефицитом CD73. Например, аденозин, генерируемый CD73, уменьшает воспаление и фиброз в легких мышей, получавших блеомицин 15 , и является толерогенным для аллотрансплантатов сердца и дыхательных путей. 16,17 CD73-зависимый A 2B Передача сигналов AR защищает мышей во время почечной ишемии, 18 ингибирует системную утечку из сосудов во время гипоксии, 19,20 и также необходима для кардиопротекции в результате ишемического прекондиционирования. 21 Внеклеточный аденозин ингибирует активацию тромбоцитов и адгезию лейкоцитов к эндотелию сосудов 22 и ограничивает миграцию лимфоцитов в дренирующие лимфатические узлы после воспалительного стимула посредством CD73-зависимого механизма. 23 CD73 также ингибирует противоопухолевый иммунный ответ. 24-27
Существует несколько дополнительных модельных систем, в которых было показано, что передача сигналов AR играет роль в иммунной регуляции, но важность CD73 в выработке аденозина для запуска рецепторов не изучалась. Например, была задокументирована иммунная регуляция посредством активации A 2A и/или A 2B AR в Т-клетках и дендритных клетках (DC). 28,29 Кроме того, было показано, что специфический агонист A 2A AR ингибирует аллогенную иммунную активацию in vitro посредством воздействия как на Т-клетки, так и на антигенпрезентирующие клетки, а также задерживает отторжение аллотрансплантата кожи in vivo. 14 Было показано, что другой агонист A 2A AR ослабляет острую РТПХ после алло-HCT. 13 Эти результаты подтверждают концепцию о том, что внеклеточный аденозин противодействует активации иммунной системы, запускаемой АТФ, как механизм отрицательной обратной связи, предотвращающий неконтролируемое разрушение тканей из-за чрезмерного воспаления.
Например, была задокументирована иммунная регуляция посредством активации A 2A и/или A 2B AR в Т-клетках и дендритных клетках (DC). 28,29 Кроме того, было показано, что специфический агонист A 2A AR ингибирует аллогенную иммунную активацию in vitro посредством воздействия как на Т-клетки, так и на антигенпрезентирующие клетки, а также задерживает отторжение аллотрансплантата кожи in vivo. 14 Было показано, что другой агонист A 2A AR ослабляет острую РТПХ после алло-HCT. 13 Эти результаты подтверждают концепцию о том, что внеклеточный аденозин противодействует активации иммунной системы, запускаемой АТФ, как механизм отрицательной обратной связи, предотвращающий неконтролируемое разрушение тканей из-за чрезмерного воспаления.
Цель этого исследования состояла в том, чтобы определить роль CD73 в ответах на острую РТПХ и трансплантат против лейкемии (GVL).