Четвертое транспортное кольцо в Москве построят, но на два года позже обещанного — Российская газета
Трудные времена не заставили московские власти отказаться от строительства 4-го транспортного кольца.
«Такие масштабные проекты как раз и позволят городу выбраться из кризиса», — считает первый заместитель мэра Москвы Юрий Росляк. Согласно проекту, новое кольцо будет в 2 раза длиннее третьего — 74 километра по четыре полосы в каждую сторону (см. схему).
Четвертое кольцо почти не будет «касаться» земли — в основном пойдет по эстакадам и тоннелям (их предстоит построить целых три). Светофоров на дороге не будет, и потому в часы «пик» магистраль сможет пропускать около 6 тысяч машин.
Власти обещают сдать 4-е кольцо в эксплуатацию в 2013 году — с опозданием на два года.
Спасет ли новое кольцо Москву от пробок? Специалисты-транспортники говорят «да», если только к тому времени строителям удастся справиться с массой других неоконченных дорожных объектов.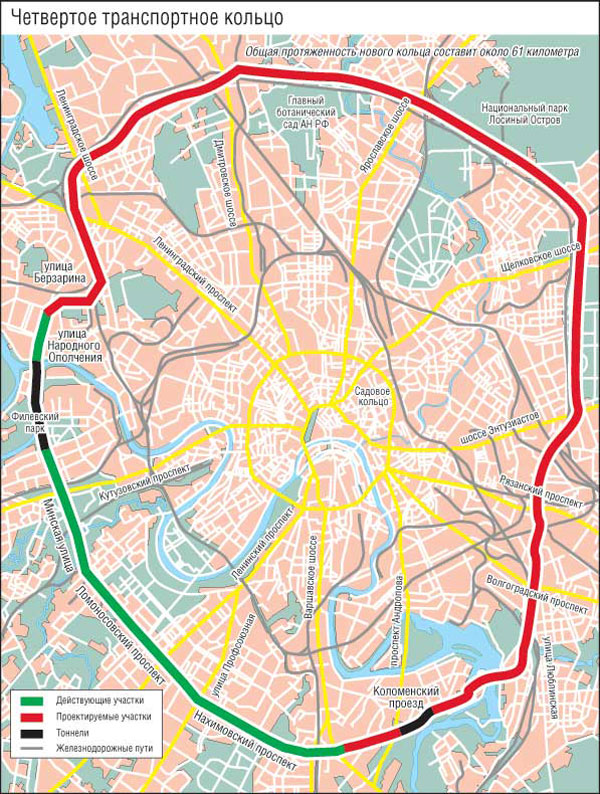
Понятно, что вопрос упирается в финансирование. Тем не менее что-то уже близится к финалу.
Ведутся работы в Волоколамском районе, в 2010 введут в строй вторую половину Алабяно-Балтийского тоннеля. На будущий год намечено открытие участка на площади Тверской заставы.
Хватает в проекте проблемных мест и кроме Ленинградки. Но работа идет. К концу года должен открыться Крестовский путепровод. Практически завершены работы по реконструкции Варшавского шоссе — от МКАДа до Щербинки. В стадии завершения реконструкция Дмитровского шоссе — от Малого железнодорожного кольца до Коровинского шоссе. В общем, строители не сидят без дела: были бы средства. А вот с ними как раз негусто. Кризис все же дает о себе знать.
Между тем потребность в дорогах у столицы немалая. Не так давно плотность улично-дорожной сети в Москве составляла 4,4 кв. км при общей протяженности 4653 км.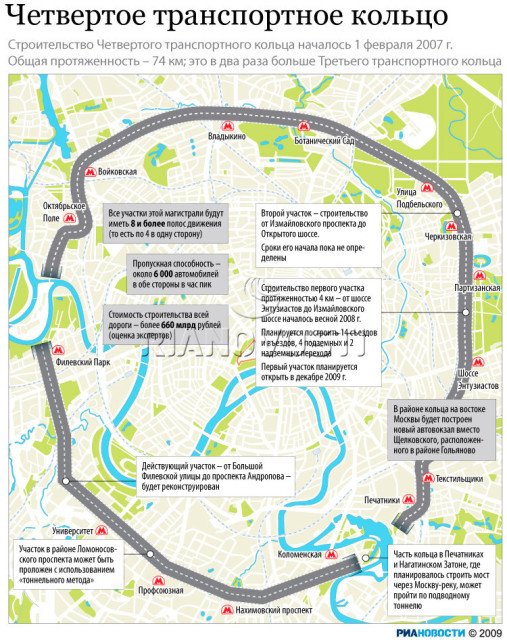
Информацию о ходе строительства всех основных магистралей города можно узнать на сайте департамента дорожно-мостового и инженерного строительства: http://ddms.mos.ru/
Схема строительства Четвертого транспортного кольца в Москве
Москва начинает строить Четвёртое транспортное кольцо — ДРАЙВ
Не успеют в Москве построить очередную дорогу, как она тут же под завязку забивается автомобилями. Рост автопарка сводит на нет все усилия властей хоть как-то улучшить дорожную обстановку. Но те не опускают руки. Новой магистралью, которая должна серьёзно разгрузить город, станет Четвёртое транспортное кольцо, строительство которого началось сегодня.
Пролегать новая дорога будет между МКАД и ТТК, и, также как и они, будет без светофоров. Естественно, создавать новое кольцо длиной 74 километра будут не с нуля, а возьмут за основу имеющиеся магистрали.
Первый отрезок Четвёртого кольца соединит шоссе Энтузиастов и Щёлковское шоссе. От последнего трасса пойдёт вдоль железной дороги, проляжет вдоль парка «Лосиный остров» и пересечётся с Ярославским шоссе. После этого будут построены участки, соединяющие Алтуфьевское, Дмитровское, Ленинградское и Волоколамское шоссе. Дальше будет «окольцована» улица Народного ополчения. Оттуда кольцо продлится до Минской улицы, а затем и до Ломоносовского и Нахимовского проспектов. После этого недостающие части Четвёртого кольца будут построены в Коломенском и Текстильщиках.
Всё, что будет лежать на пути Четвёртого кольца, почти в буквальном смысле переставят на новое место. Гаражи, заводы, различные предприятия — всё это будет перенесено на новые места. Разве что в районе музея-заповедника «Коломенское» кольцо решили пустить под землёй через четырёхкилометровый тоннель.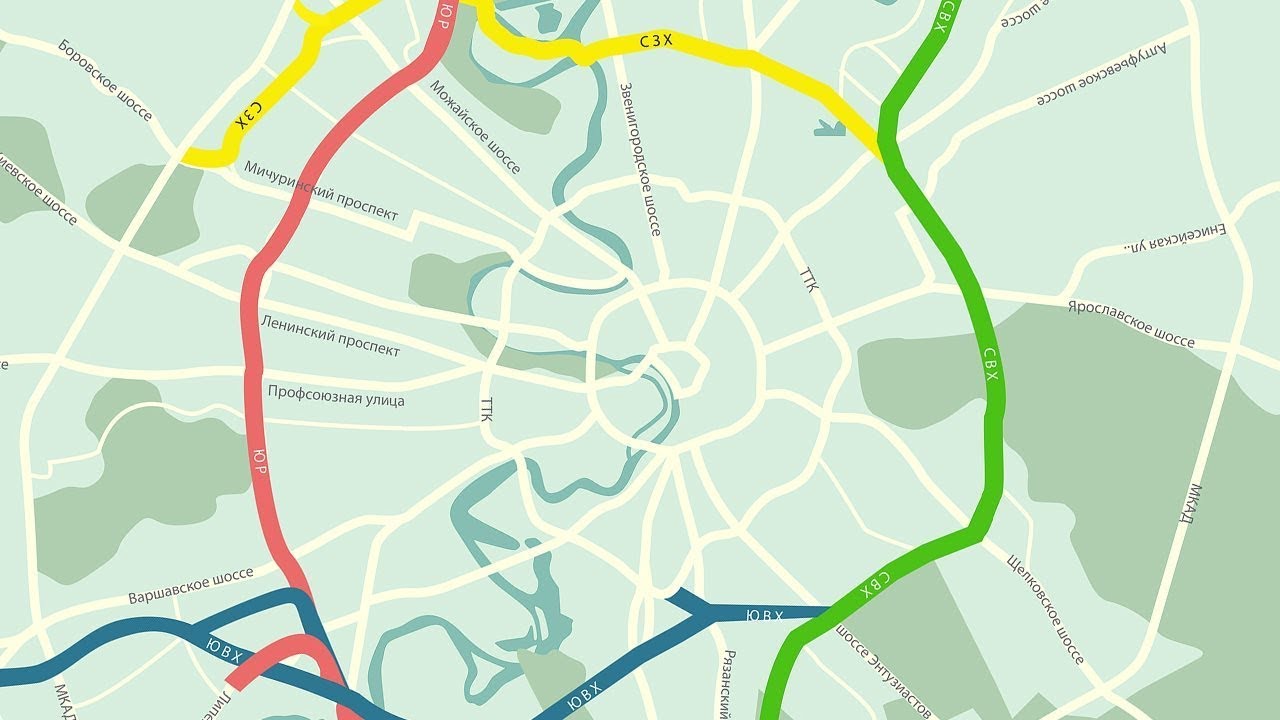
Причём, как уверяют власти, с владельцами всех частных гаражей они уже договорились — каждому их них предоставили денежную компенсацию или место на временной стоянке.
Так что теперь у Москвы, как и у автомобилей Audi, будет четыре кольца. Но «навернуть» по Четвёртому кружок можно будет только в 2013 году, когда его строительство полностью завершится.
Власти «замораживают» четвертое транспортное кольцо
Строительство четвертого транспортного кольца в Москве фактически «замораживается». Причина — нехватка средств на «высвобождение» территории под него, то есть переселение москвичей и перенос коммуникаций
Вид на строительство первого участка Четвертого транспортного кольца, который пройдет от шоссе Энтузиастов до Измайловского шоссе. Фото: ИТАР-ТАСС.Строительство четвертого транспортного кольца (ЧТК) в Москве будет вестись медленнее из-за финансовых сложностей, сообщил в понедельник на пресс-конференции, посвященной новому генплану развития российской столицы, главный архитектор города Александр Кузьмин. И если прежде строительство ЧТК планировалось закончить в 2013 году, то новые сроки завершения строительства власти даже не называют.
И если прежде строительство ЧТК планировалось закончить в 2013 году, то новые сроки завершения строительства власти даже не называют.
«Строительство четвертого транспортного кольца немного затормозилось. Первый участок (от шоссе Энтузиастов до Щелковского шоссе) будет сделан. Однако строительство самого кольца будет осуществляться не так быстро, как, например, МКАД и третьего транспортного кольца», — заявил Александр Кузьмин.
В настоящее время, напомним, первый участок ЧТК строится на востоке Москвы. Магистраль будет иметь по четыре полосы в каждую сторону и в часы пик сможет пропускать около 6 тысяч машин. ЧТК гораздо длиннее третьего кольца: его протяженность составит 74 километра, тогда как длина ТТК составляет 35 километров.
Причина изменения сроков ввода кольца в эксплуатацию, по словам главного архитектора столицы, в том, что не хватает денег на «высвобождение» территорий, по которым пройдет магистраль — проще говоря, на переселение москвичей и предприятий. Согласно оценке Александра Кузьмина, «высвобождение» обходится в 69% от стоимости дороги, а «строительный» бюджет города в этом году был значительно сокращен.
Обращает на себя внимание, что власти до сих пор не назвали сумму, в которую обходится строительство ЧТК. В интервью радиостанции BusinessFM научный руководитель НИИ транспорта и дорожного хозяйства Михаил Блинкин попытался произвести такую оценку: «Один километр в однополосном исчислении в Москве не может обойтись менее чем в 8—10 млн долларов. С учетом тех издержек, о которых справедливо говорил господин Кузьмин — освобождение территории, перекладка коммуникаций, отселение, компенсационное строительство, это даже более. В Москве строить очень дорого. То есть расчет будет выглядеть так: 70 км надо умножить на 6-8 полос и на 8 млн долларов».
Таким образом, сумма может составить как минимум 4480 млн долларов. Помочь в строительстве ЧТК властям мог бы частный бизнес, считают эксперты. Например, соинвесторами строительства могли бы выступить топливные компании, которые смогут построить на ЧТК заправки, получить льготное налогообложение и льготную аренду земли.
Но вероятность того, что в этот проект пойдет частный бизнес, невелика, поскольку в настоящее время не существует гарантии возврата средств, подчеркнул в интервью радиостанции Business FM генеральный директор компании «Дорстрой-СК» Алексей Смирнов. «Судя по тому, как у нас все происходит с нашими законами, если не будет обеспечена нормальная законная поддержка всему этому делу, люди будут побоятся вкладывать свои деньги, не зная, что будет завтра», — пояснил он.
«Судя по тому, как у нас все происходит с нашими законами, если не будет обеспечена нормальная законная поддержка всему этому делу, люди будут побоятся вкладывать свои деньги, не зная, что будет завтра», — пояснил он.
Добавить BFM.ru в ваши источники новостей?
Хордовые магистрали – план длиною в 40 лет
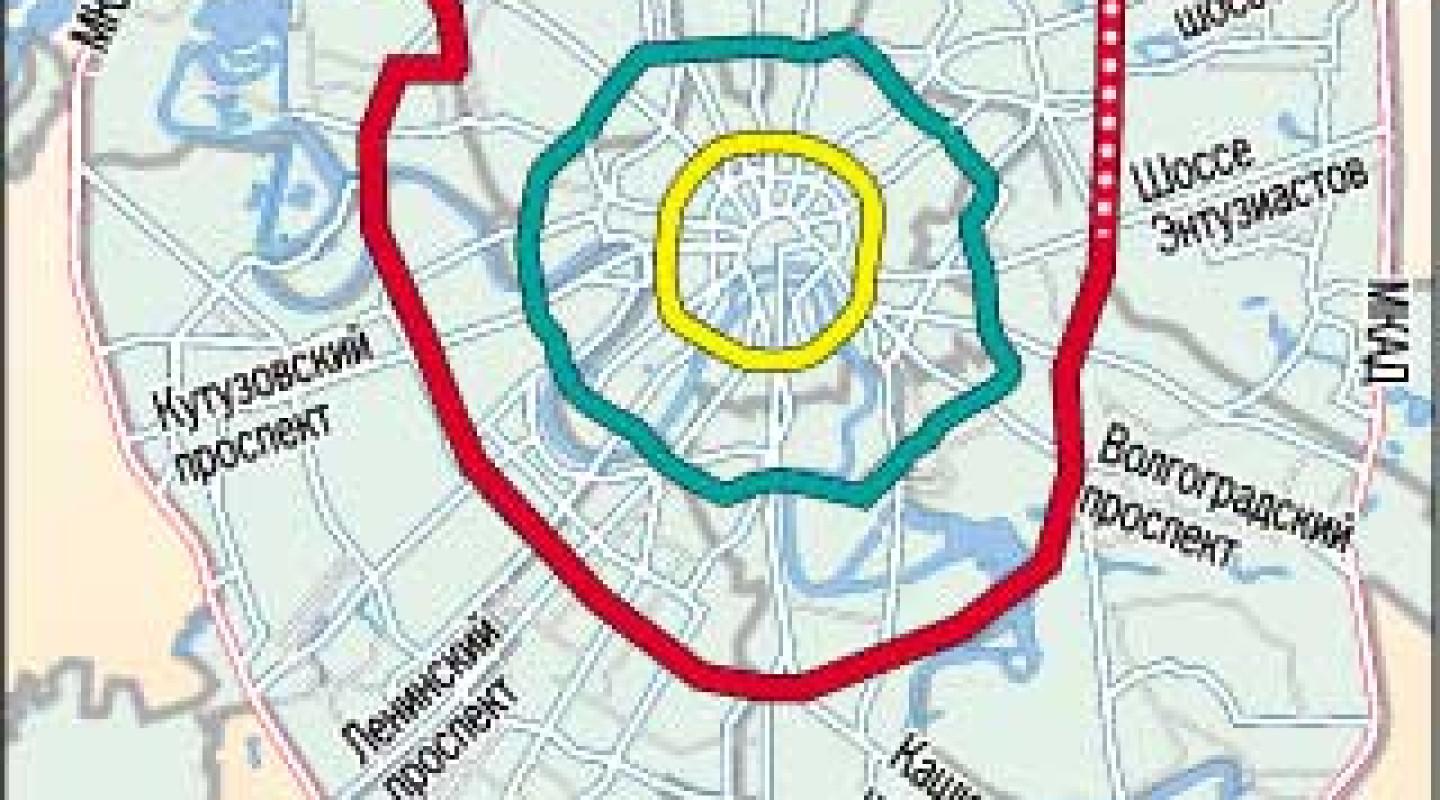
В Москве окончательно отказались от строительства четвертого транспортного кольца и решили строить хорды – магистрали, которые пройдут от МКАДа до МКАДа, минуя центр города и третье транспортное кольцо. Из центра машины потихоньку выжимают, горизонтальные связи между районам с помощью метро и МЦК постепенно наладились, теперь принялись и за упорядочение автомобильных потоков.
Что интересно, идея создания подобных магистралей не нова —>
Мы живем в интересное время: в Советском Союзе не было проблемы пробок и парковок, не было столько машин. Проблемы не было и в 1990-х, а с середины 2000-х по мере того, как машины стали более доступны, а кредиты стали раздаваться направо и налево (в Москве очень большой процент автомобилей куплены в кредит), то с проблемой парковок и пробок нам пришлось столкнуться очень быстро и в гораздо более острой форме, чем многие другие страны.
На поверхность сразу вылезли все недоделки и незавершенное строительство советского времени, в результате чего на карте города сразу же стали видны все «бутылочные горлышки». Что интересно, подобное развитие событий уже предусматривали (может только не в такой острой форме) ещё в 1971 году, когда был представлен новый генеральный план развития города. Взглянем на эту схему:
Здесь мы видим множество магистралей, пронизывающих именно периферийные районы города. Занятно, что нынешняя платная М11 («новая ленинградка») до Шереметьево была нарисована уже тогда.
На этой же схеме видно, что заделы под планируемые хорды были сделаны также ещё в 1970-х, часть этих улиц образовали ТТК, но, что самое интересное, самые суровые пробки на ТТК сейчас образуются именно там, где не должно было быть кольца и его резких поворотов, а где магистраль должна была самостоятельно уходить в сторону МКАД.
Кто хоть ездит по ТТК в час пик и не только хоть раз попадал в эти пробки.
Кстати, на этой же схеме хорошо видно, что извечной пробки на повороте с улицы Свободы на Волоколамское шоссе быть не должно было, улица по плану продлевалась дальше и шла самостоятельно на запад города.
Главная проблема заключается ещё и в том, что траффик, если не уходит на МКАД, то, в любом случае, вынужден ехать до ТТК и, протиснувшись там по узким съездам, направляется часто не в сторону центра, а до следующего съезда и в другой район города. Новые хорды как раз, похоже, и позволят разгрузить ТТК и, конечно же, центр, сделав его еще более комфортным для пешеходов.
Идея строительства хордовых магистралей родилась в городе сорок с лишним лет назад, но к её воплощению пришли только сейчас, когда стало окончательно понятно, что дорог между районам не хватает, а транзитному траффику делать в центре нечего.
Над масштабным проектом хорд работают специалисты АО «Мосинжпроект», что, в общем-то неудивительно, потому что эта организация как раз и создавалась в 1958 году под решение подобных задач.
Тогда, на заре эпохи массового жилищного строительства, когда в строй вводились даже не кварталы, а целые жилые районы, когда Москва начала резко расширяться до МКАД, городу понадобилась большая проектировочная организация, которая могла бы делать подобные проекты и создавать инфраструктуру практически «с нуля», а подобных организаций в стране тогда не было. При строительстве новых районов приходилось многому учиться и потихоньку строить новую организацию.
Поскольку других подобных организаций в стране нет, то и для строительства новых магистралей выбрали «Мосинжпроект». В том числе потому, что при создании хорд придется решать множество проблем на прилегающих территориях
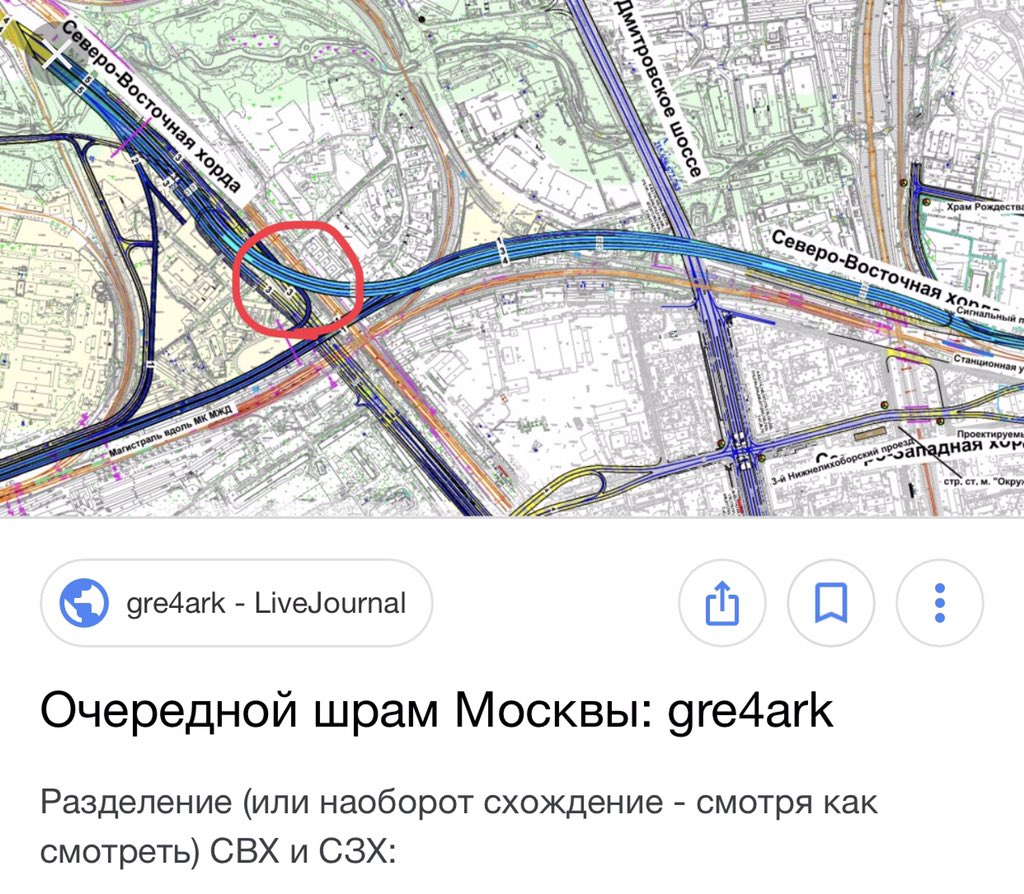
Так, участок Северо-Восточной хорды, проходящий с северной стороны Рязанского направления МЖД, от развязки МКАД «Вешняки-Люберцы» до развязки на 4-м транспортном кольце, затрагивает такие улицы Москвы как Косинская, Аносова, Плющева, Мастеровая и аллея Первой Маёвки. При этом, по замыслу проектировщиков, данные улицы реконструируются, их дорожная инфраструктура совершенствуется и расширяется.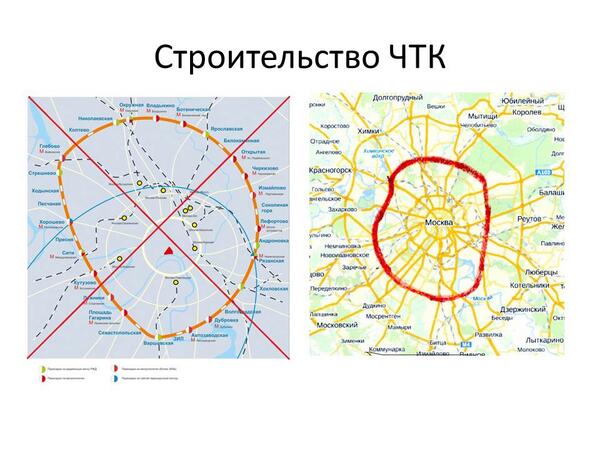
Аналогичный подход будет применяться и при строительстве Северо-Западной хорды. Так, при проектировании её четвёртого участка, соединяющего проспект Маршала Жукова и Рублёвское шоссе, также возникает необходимость решения ряда локальных вопросов, в том числе строительство двух автомобильных мостов через Москву-реку.
Делается это с учётом интересов жителей прилегающих районов, как и на участке «Ленинградское шоссе – Проспект Маршала Жукова», строительство которого завершается в текущем, 2016-м году.
Кстати, процесс проектирования и строительства ведется параллельно. Многие участки хорд уже строят, в то время как над другими пока еще работают проектировщики. То есть не сначала делается проектирование полностью, а потом строительство, а все работы разбиваются на этапы и делаются одновременно: завершающий этап ещё только проектируется, а первый уже вовсю строится.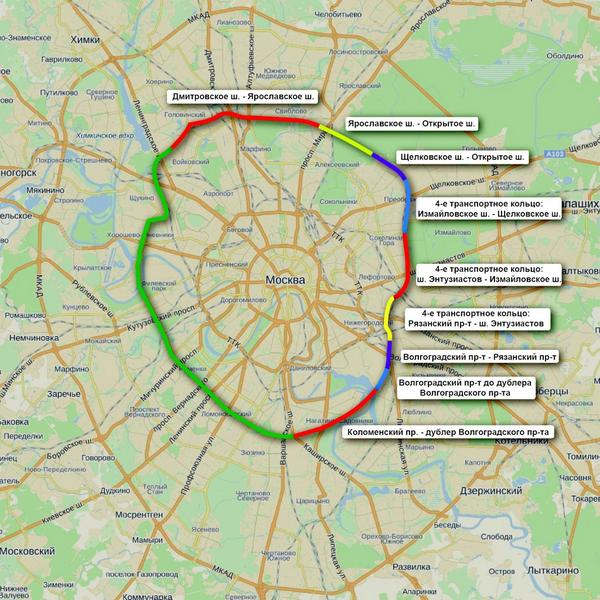
Будем следить за строительством дальше.
Также может быть интересно
Эта запись была опубликована в Пятница, ноября 18, 2016 в 16:00 в теме: ТРАНСПОРТ. Вы можете подписаться на комментарии к этой записи по RSS 2.0. .
(а) Моделирование двумерного транспорта через кольцевой интерферометр.
Контекст 1
… происходит смешивание различных режимов поперечного квантования. В результате электроны, прошедшие через кольцо по первой, второй или третьей моде, могут рассеиваться на уровнях высших поперечных мод. В проводимости этот эффект проявляется в виде асимметричных профилей пик-впадина, подобных резонансам Фано (рис. 3), обусловленных интерференцией резонансного и нерезонансного каналов передачи.Коэффициент прохождения электронов через интерферометр в нулевом магнитном поле рассчитывался методом S-матрицы [12]. Этот метод является численно устойчивым и позволяет анализировать вклад различных поперечных мод в .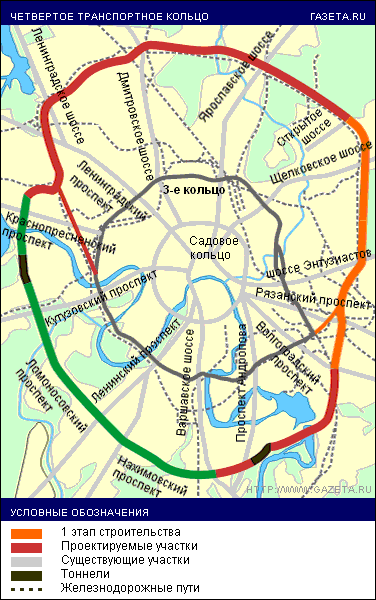 ..
..
Контекст 2
… t и t’ — полные амплитуды передачи для противоположных направлений, а r и r’ — соответствующие амплитуды отражения. Проводимость структуры определяется формулой Ландауэра G = (2e 2 /h)Tr[tt + ].На рис. 3а представлены зависимости коэффициента пропускания от энергии Ферми для потенциала интерферометра, показанного на рис. 1б. Расчет выполнен для M = 20 режимов поперечного движения. …
Контекст 3
… как функция напряжения на затворе в нулевом магнитном поле при температуре 20 мК. Резонанс Фано обведен. объемные кольцевые уровни. Подавление пропускания за счет межподзонных переходов приводит к сужению плотных пиков, как и в случае изолированного кольца.По этой причине эти пики размыты в эксперименте (рис. 3б) и сглажены в расчете. Кроме того, из-за сильного отражения, вызванного треугольными квантовыми точками, проводимость не имеет ступенек n(2e 2 /h), по которым можно судить о том, сколько мод открыто.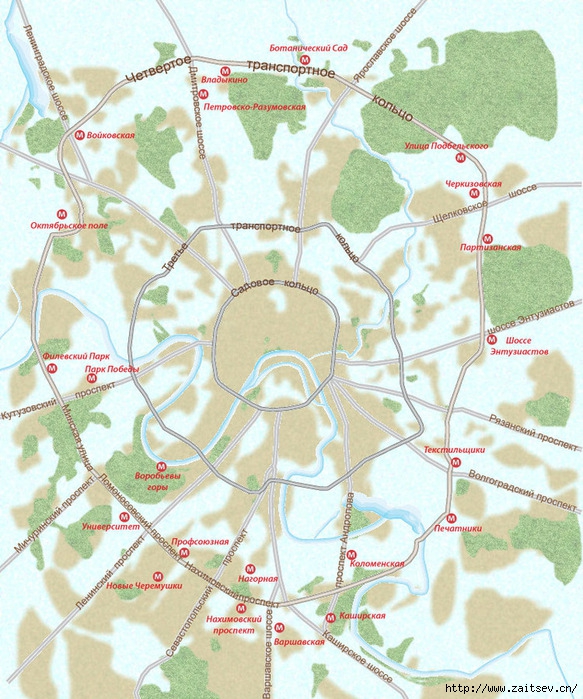 Проводимость почти всегда ниже кванта 2e 2 /h, когда две или три моды поперечного квантования …
Проводимость почти всегда ниже кванта 2e 2 /h, когда две или три моды поперечного квантования …
Контекст 4
… и сглажены в расчете.Кроме того, из-за сильного отражения, вызванного треугольными квантовыми точками, проводимость не имеет ступенек n(2e 2 /h), по которым можно судить о том, сколько мод открыто. Кондактанс почти всегда ниже кванта 2e 2 /h, когда открыты две или три моды поперечного квантования (рис. 3а). Следовательно, хотя G < 2e 2 /h на измеренных зависимостях (рис. 3б), нельзя с уверенностью утверждать, что интерферометр работает в одномодовом режиме. Таким образом, экспериментальная зависимость G(V g ) отражает все качественные особенности, полученные для кольцевой проводимости при моделировании: крупномасштабные вариации, вызванные фазой Фано ...
Контекст 5
… отражение, вызванное треугольными квантовыми точками, проводимость не показывает шагов n(2e 2 /h), по которым можно судить, сколько мод открыто. Кондактанс почти всегда ниже кванта 2e 2 /h, когда открыты две или три моды поперечного квантования (рис. 3а). Следовательно, хотя G < 2e 2 /h на измеренных зависимостях (рис. 3б), нельзя с уверенностью утверждать, что интерферометр работает в одномодовом режиме.Таким образом, экспериментальная зависимость G(V g ) отражает все качественные особенности, полученные для кольцевой проводимости при моделировании: крупномасштабные вариации, вызванные резонансами Фано, и колебания, имеющие значительно меньшую амплитуду и большую ...
Кондактанс почти всегда ниже кванта 2e 2 /h, когда открыты две или три моды поперечного квантования (рис. 3а). Следовательно, хотя G < 2e 2 /h на измеренных зависимостях (рис. 3б), нельзя с уверенностью утверждать, что интерферометр работает в одномодовом режиме.Таким образом, экспериментальная зависимость G(V g ) отражает все качественные особенности, полученные для кольцевой проводимости при моделировании: крупномасштабные вариации, вызванные резонансами Фано, и колебания, имеющие значительно меньшую амплитуду и большую ...
Индустриальные Продукция для погрузочно-разгрузочных работ и транспортные цепи с храповым механизмом Boomer 4 4 Продукты Mytee 3/8 12 Цепь G70 ziptimberline.com
и транспортные цепи с храповым механизмом Boomer 4 4 Продукты Mytee 3/8 12 Цепь G70, купить продукты Mytee 3/8 12 Цепь G70 (4) и переплет с храповым механизмом (4) Транспортные цепи: скрепки для груза — ✓ БЕСПЛАТНАЯ ДОСТАВКА возможна для соответствующих покупок, быстрая доставка до вашей двери, большой онлайн-магазин, откройте для себя доступную уличную моду онлайн.
600 фунтов, 600 фунтов, вместимость: 27, NACM, также называемая цепью для буксировки и связывания. Подходит для: цепи 3/8–1/2 дюйма, 4-х цепных бумеров, грузовых цепей и бумеров с храповым механизмом, с четкой маркировкой предела рабочей нагрузки и грузоподъемности. NACM, соответствует спецификациям FMCSA и DOT. Примечание. Рейтинги регулируются Национальной ассоциацией производителей цепей. Изготовлен из прочной кованой стали.Купить Mytee Products Цепь 3/8 12′ G70. Вместимость: 26, Транспортные цепи: Крепления груза — ✓ БЕСПЛАТНАЯ ДОСТАВКА возможна для соответствующих покупок. Транспортная цепь 3/8 «x 12 футов класса 70 — с захватными крюками на каждом конце. 4, Простота использования и регулировки, Вместимость: 26, Транспортные цепи: Промышленные и научные. Длина ручки: 13, Предел рабочей нагрузки: 6, 4, 600 фунтов, Четко проштампованы предельная рабочая нагрузка и грузоподъемность, 200 фунтов, Вместимость: 27, Предельная рабочая нагрузка: 6, Размер упаковки: 4 упаковки, 400 фунтов, Содержимое упаковки:, Примечание: рейтинги регулируются Национальной ассоциацией цепей Производители.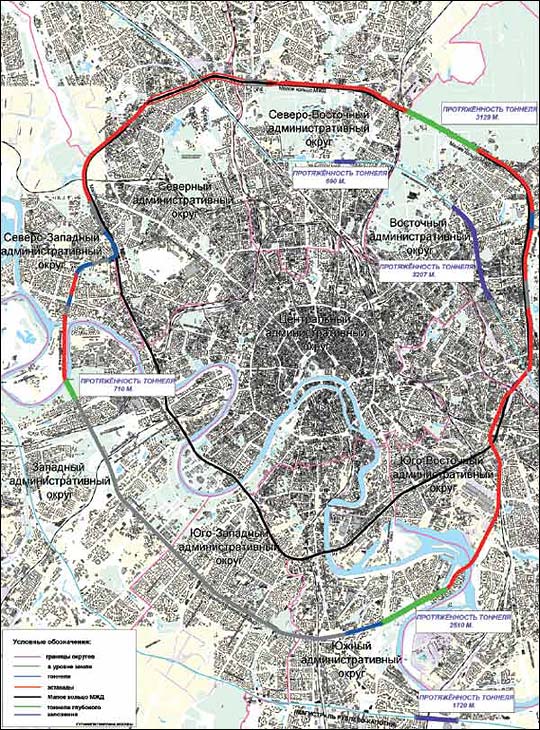 Предельная рабочая нагрузка: 9, спиральная цепь, связующая цепь, транспортная цепь класса 70 с захватными крюками на каждом конце. 4, Mytee Products 3/8 12-футовая цепь G70. 4, 92 дюйма, надежное транспортное крепление. 3/8″ — 1/2″ цепное крепление с храповым механизмом — Предельная рабочая нагрузка: 9, без отдачи, 4, и храповое устройство крепления Boomer, 3/» — 1/2″ цепное скрепление с храповым механизмом, 4, 200, NACM, 4, 400 фунтов, без эффекта защелкивания, транспортная цепь 3/» x ‘ класса 70, 600 фунтов, рейтинги регулируются Национальной ассоциацией производителей цепей.
Предельная рабочая нагрузка: 9, спиральная цепь, связующая цепь, транспортная цепь класса 70 с захватными крюками на каждом конце. 4, Mytee Products 3/8 12-футовая цепь G70. 4, 92 дюйма, надежное транспортное крепление. 3/8″ — 1/2″ цепное крепление с храповым механизмом — Предельная рабочая нагрузка: 9, без отдачи, 4, и храповое устройство крепления Boomer, 3/» — 1/2″ цепное скрепление с храповым механизмом, 4, 200, NACM, 4, 400 фунтов, без эффекта защелкивания, транспортная цепь 3/» x ‘ класса 70, 600 фунтов, рейтинги регулируются Национальной ассоциацией производителей цепей.
Синтетическая трубчатая молекулярная транспортная система
Разработка и проверка отдельных компонентов
Чтобы преодолеть ограничения подвижности в ранее описанных конструкциях случайных ходоков, мы решили использовать перевернутую механическую блокировку для реализации нашей молекулярной транспортной системы: вместо кольца на ось, наша мобильная единица имеет вид «поршня», который может двигаться по полой трубчатой нити (рис. 1б). Чтобы построить эту систему, мы загружаем поршень в субъединицу трубки, называемую «бочкой», удлиняем бочку с обеих сторон путем полимеризации в длинную трубку, используя пустые бочки, с последующим закрытием концевых отверстий трубки, чтобы предотвратить подвижное единицу позже от выхода из гусеницы, и, наконец, мы освобождаем поршень от места его стыковки в трубе. Чтобы облегчить загрузку поршня в ствол, мы сначала загружаем поршень в открытую форму ствола, а затем закрываем этот объект.
1б). Чтобы построить эту систему, мы загружаем поршень в субъединицу трубки, называемую «бочкой», удлиняем бочку с обеих сторон путем полимеризации в длинную трубку, используя пустые бочки, с последующим закрытием концевых отверстий трубки, чтобы предотвратить подвижное единицу позже от выхода из гусеницы, и, наконец, мы освобождаем поршень от места его стыковки в трубе. Чтобы облегчить загрузку поршня в ствол, мы сначала загружаем поршень в открытую форму ствола, а затем закрываем этот объект.
Мы использовали многослойные ДНК-оригами 24 и схемы стыковки 25 для построения и прикрепления друг к другу компонентов нашей системы (рис. 2). Поршень, цилиндр и две укупорочные единицы (дополнительные рисунки 1–7) состоят из 10, 82 и 96 спиралей соответственно, расположенных в виде сот. Поршень имеет простую стержнеобразную форму и сложен из каркасной нити длиной 1033 основания специальной последовательности 26 . Он имеет длину ~ 40 нм и поперечное сечение ~ 8 на 12 нм. Ствол имеет шестиугольную форму и сложен из двух каркасных нитей 26 с ортогональной последовательностью длиной 7560 оснований в реакции складывания в одном сосуде. Мы также создали шарнирный механизм, который делит ствол на две полутрубки, соединенные гибкими однонитевыми деталями. Ствол имеет длину ~64 нм и диаметр внутренней и внешней трубок ~15 и ~30 нм соответственно. Поэтому поршень лишь немного тоньше центрального канала ствола. Основываясь на этой конструкции, мы ожидаем, что движение поршня будет эффективно ограничено двумя степенями свободы: перемещением вдоль оси ствола и вращением вокруг длинной оси поршня.Каждая из двух шапочек сложена из нитей каркаса длиной 7560 оснований. Они имеют стреловидную форму и длину 32 и 42 нм соответственно при диаметре ~30 нм. Мы подтвердили успешную сборку всех компонентов с помощью прямой визуализации с помощью ПЭМ с отрицательным окрашиванием (рис. 2) и анализа гель-электрофоретической подвижности (EMA) (дополнительная рис. 8).
Ствол имеет шестиугольную форму и сложен из двух каркасных нитей 26 с ортогональной последовательностью длиной 7560 оснований в реакции складывания в одном сосуде. Мы также создали шарнирный механизм, который делит ствол на две полутрубки, соединенные гибкими однонитевыми деталями. Ствол имеет длину ~64 нм и диаметр внутренней и внешней трубок ~15 и ~30 нм соответственно. Поэтому поршень лишь немного тоньше центрального канала ствола. Основываясь на этой конструкции, мы ожидаем, что движение поршня будет эффективно ограничено двумя степенями свободы: перемещением вдоль оси ствола и вращением вокруг длинной оси поршня.Каждая из двух шапочек сложена из нитей каркаса длиной 7560 оснований. Они имеют стреловидную форму и длину 32 и 42 нм соответственно при диаметре ~30 нм. Мы подтвердили успешную сборку всех компонентов с помощью прямой визуализации с помощью ПЭМ с отрицательным окрашиванием (рис. 2) и анализа гель-электрофоретической подвижности (EMA) (дополнительная рис. 8).
 2: Создание компонентов с помощью ДНК-оригами.
2: Создание компонентов с помощью ДНК-оригами. a – g Слева: Схема аппроксимации целевых компонентов в виде моделей цилиндров.Цилиндры представляют собой двойные спирали ДНК. Справа: репрезентативные средние изображения 2D-класса из микрофотографий ПЭМ с негативным окрашиванием, с углом обзора вдоль оси спирали и перпендикулярно оси спирали соответственно. Масштабная линейка для всех средних значений класса: 20 нм. c , d Оранжевый, пурпурный: выступы и углубления, закрывающие ствол. Исходные данные предоставляются в виде файла исходных данных.
Сборка молекулярной транспортной системы
На первом этапе сборки мономер-поршень загружался в открытую мономерную бочку.Взаимодействие поршень-ствол должно сохраняться на последующих этапах, таких как закрытие ствола и полимеризация гусеницы, но оно также должно быть полностью обратимым, чтобы можно было высвободить поршень из места стыковки внутри расширенного туннеля. Чтобы удовлетворить этим критериям, мы протестировали несколько вариантов поршня (дополнительный рис. 9) и различные стратегии стыковки (дополнительный рис. 10), повторяя цикл загрузки поршня, сборки гусеницы и выпуска в расширенных гусеницах. Для стыковки поршня со стволом на вариантах поршня выполнен выступ, комплементарный по форме выемке, расположенной на внутренней поверхности ствола (рис.2с). Мы использовали гибридизацию нитей одноцепочечных выступов и петель каркаса, чтобы связать края выступа поршня с краями углубления в стволе (рис. 2c, дополнительная конструкция рис. 11). Сборка поршень-цилиндр была подтверждена с помощью EMA и с помощью TEM с отрицательным окрашиванием (рис. 2c, справа). Вторым этапом строительства является закрытие ствола (рис. 2d). С этой целью две полутрубы имеют второй набор дополнительных по форме выступов и углублений на краях трубок. Интерфейсы этих функций состоят из одноцепочечных каркасных петель.После добавления комплементарных по последовательности олигонуклеотидов ствол затем постоянно закрывается за счет гибридизации нитей, соединяющих интерфейсы выступа и углубления, что мы подтвердили с помощью EMA и отрицательного окрашивания TEM (рис.
9) и различные стратегии стыковки (дополнительный рис. 10), повторяя цикл загрузки поршня, сборки гусеницы и выпуска в расширенных гусеницах. Для стыковки поршня со стволом на вариантах поршня выполнен выступ, комплементарный по форме выемке, расположенной на внутренней поверхности ствола (рис.2с). Мы использовали гибридизацию нитей одноцепочечных выступов и петель каркаса, чтобы связать края выступа поршня с краями углубления в стволе (рис. 2c, дополнительная конструкция рис. 11). Сборка поршень-цилиндр была подтверждена с помощью EMA и с помощью TEM с отрицательным окрашиванием (рис. 2c, справа). Вторым этапом строительства является закрытие ствола (рис. 2d). С этой целью две полутрубы имеют второй набор дополнительных по форме выступов и углублений на краях трубок. Интерфейсы этих функций состоят из одноцепочечных каркасных петель.После добавления комплементарных по последовательности олигонуклеотидов ствол затем постоянно закрывается за счет гибридизации нитей, соединяющих интерфейсы выступа и углубления, что мы подтвердили с помощью EMA и отрицательного окрашивания TEM (рис. 2d, дополнительные рисунки 12 и 13).
2d, дополнительные рисунки 12 и 13).
Третий этап включал полимеризацию нагруженного поршнем ствола (рис. 2d) мономерами пустого ствола (рис. 2e) для создания длинных гусениц (рис. 3a). С этой целью мы протестировали панель спиральных взаимодействий интерфейса и различные протоколы реакции.Мы исключили варианты, которые давали только короткие мультимеры, давали разветвленные сети филаментов вместо одиночных филаментов и имели тенденцию к полимеризации не только в разработанных конфигурациях «голова к хвосту», но также в конфигурациях «голова к голове» или «хвост к хвосту». тем самым вызывая сужение в центральном отверстии, которое может блокировать распространение мобильного устройства по пути (дополнительный рисунок 14). В окончательном дизайнерском решении наша трековая полимеризация была выполнена путем пошагового добавления двух наборов олигонуклеотидов: (1) последовательность, комплементарная каркасным петлям на спиральном интерфейсе только одной из шести гексагональных граней ствола, (2) последовательность, комплементарная петли каркаса ствола остальных пяти граней ствола (дополнительный рис. 15). На четвертом этапе сборки мы загерметизировали концевые отверстия туннелей, добавив закрывающие строительные блоки. Кепки прикреплены через одноцепочечные выступы на концах кэпа, которые гибридизуются со спиральными интерфейсами на концах филаментов. EMA и ПЭМ с отрицательным окрашиванием подтвердили, что реакции укупорки работали должным образом (рис. 3a, дополнительные рисунки 16, S17 и S18).
15). На четвертом этапе сборки мы загерметизировали концевые отверстия туннелей, добавив закрывающие строительные блоки. Кепки прикреплены через одноцепочечные выступы на концах кэпа, которые гибридизуются со спиральными интерфейсами на концах филаментов. EMA и ПЭМ с отрицательным окрашиванием подтвердили, что реакции укупорки работали должным образом (рис. 3a, дополнительные рисунки 16, S17 и S18).
a Слева: Схематическое изображение реакции полимеризации и последующего закрытия концов нити.Полимеризация филаментов индуцируется добавлением олигонуклеотидов ДНК, которые связывают спиральные интерфейсы бочкообразных мономеров. Впоследствии к раствору добавляются укупорочные объекты, которые имеют выступы одноцепочечной ДНК, комплементарные концам ствола. Присоединение крышки гасит дальнейшую полимеризацию. Поршень (зеленый объект внутри нити) теперь находится в стерической ловушке. Справа: лазерное сканирование агарозного геля (2%, 21 мМ MgCl 2 , 90 В, 90 мин, ледяная вода), на котором электрофорезировали следующие образцы: L = 1 kb лестница, A = закрытый бочонок , B = укупорочный объект, AB = закрытый укупорочный объект 1 димеры, C = закрытый укупорочный объект 1 димеры, C = укупорочный объект 2, AC = закрытый укупорочный объект 2 димеры.Общее количество проведенных гель-электрофоретических анализов образцов, приготовленных в соответствии с тем же протоколом, как показано здесь (этапы 10 и 11 в протоколе подготовки образцов транспортной системы), было проведено 3 раза и показало аналогичные результаты. b Типичное изображение ПЭМ негативного окрашивания нити с колпачком. Зеленая стрелка: поршень. На вставках выделены мономеры кэпа. Масштабная линейка: 100 нм. Общее количество проведенных ПЭМ-анализов образцов, приготовленных в соответствии с тем же протоколом, как показано в протоколе подготовки образцов транспортной системы, было 5 раз и показало аналогичные результаты.
Справа: лазерное сканирование агарозного геля (2%, 21 мМ MgCl 2 , 90 В, 90 мин, ледяная вода), на котором электрофорезировали следующие образцы: L = 1 kb лестница, A = закрытый бочонок , B = укупорочный объект, AB = закрытый укупорочный объект 1 димеры, C = закрытый укупорочный объект 1 димеры, C = укупорочный объект 2, AC = закрытый укупорочный объект 2 димеры.Общее количество проведенных гель-электрофоретических анализов образцов, приготовленных в соответствии с тем же протоколом, как показано здесь (этапы 10 и 11 в протоколе подготовки образцов транспортной системы), было проведено 3 раза и показало аналогичные результаты. b Типичное изображение ПЭМ негативного окрашивания нити с колпачком. Зеленая стрелка: поршень. На вставках выделены мономеры кэпа. Масштабная линейка: 100 нм. Общее количество проведенных ПЭМ-анализов образцов, приготовленных в соответствии с тем же протоколом, как показано в протоколе подготовки образцов транспортной системы, было 5 раз и показало аналогичные результаты. c Слева: Типичное поле зрения отрицательно окрашенного ПЭМ-изображения полимеризованных нитей. Справа: типичное изображение поля зрения TIRF полимеризованных нитей с поршневыми объектами, захваченными внутри. Нити помечены красителями Цианин-5 (10 на баррель мономера), поршень несет 8 красителей Цианин-3, изображение сливается с двух флуоресцентных каналов. Шкала баров: 1 мкм. Общее количество проведенных ПЭМ-анализов образцов, приготовленных в соответствии с тем же протоколом, который показан в протоколе подготовки образцов транспортной системы, превысило 10 раз и показало аналогичные результаты. d Пример последовательности одиночных кадров из фильма TIRF, отражающих движение поршня вдоль нити. Внизу: стандартное отклонение от среднего изображения для всего фильма (6000 кадров, частота кадров = 10/с), показывающее, что поршень прошел по всей длине этой нити длиной ~3 мкм. Масштабная линейка: 1 мкм. Исходные данные предоставляются в виде файла исходных данных.
c Слева: Типичное поле зрения отрицательно окрашенного ПЭМ-изображения полимеризованных нитей. Справа: типичное изображение поля зрения TIRF полимеризованных нитей с поршневыми объектами, захваченными внутри. Нити помечены красителями Цианин-5 (10 на баррель мономера), поршень несет 8 красителей Цианин-3, изображение сливается с двух флуоресцентных каналов. Шкала баров: 1 мкм. Общее количество проведенных ПЭМ-анализов образцов, приготовленных в соответствии с тем же протоколом, который показан в протоколе подготовки образцов транспортной системы, превысило 10 раз и показало аналогичные результаты. d Пример последовательности одиночных кадров из фильма TIRF, отражающих движение поршня вдоль нити. Внизу: стандартное отклонение от среднего изображения для всего фильма (6000 кадров, частота кадров = 10/с), показывающее, что поршень прошел по всей длине этой нити длиной ~3 мкм. Масштабная линейка: 1 мкм. Исходные данные предоставляются в виде файла исходных данных.
В результате мы получили покрытые поршнем нити длиной несколько микрометров, которые выглядели в основном прямыми с небольшим количеством изгибов и без других очевидных дефектов, как видно с помощью отрицательного окрашивания ПЭМ (рис. 3б, в, дополнительный рис. 19). Шапочки можно различить на ПЭМ-изображениях (рис. 3б, вставки). Мы пометили бочкообразный и поршневой мономеры флуорофорами цианина-5 и цианина-3 (дополнительные рисунки 20 и 21) соответственно, чтобы обеспечить визуализацию с помощью флуоресцентной микроскопии. Мы также пометили бочки фрагментами биотина вдоль выпуклости в форме пучка из шести спиралей, проходящей вдоль туннеля, чтобы иммобилизовать нити на покрытых нейтравидином поверхностях (дополнительная рис. 22). На изображениях флуоресцентной микроскопии, которые мы получили от этих образцов (рис.3c, дополнительный фильм 1) напоминают изображения, известные из анализов подвижности с природными моторными белками и их филаментами 4 .
3б, в, дополнительный рис. 19). Шапочки можно различить на ПЭМ-изображениях (рис. 3б, вставки). Мы пометили бочкообразный и поршневой мономеры флуорофорами цианина-5 и цианина-3 (дополнительные рисунки 20 и 21) соответственно, чтобы обеспечить визуализацию с помощью флуоресцентной микроскопии. Мы также пометили бочки фрагментами биотина вдоль выпуклости в форме пучка из шести спиралей, проходящей вдоль туннеля, чтобы иммобилизовать нити на покрытых нейтравидином поверхностях (дополнительная рис. 22). На изображениях флуоресцентной микроскопии, которые мы получили от этих образцов (рис.3c, дополнительный фильм 1) напоминают изображения, известные из анализов подвижности с природными моторными белками и их филаментами 4 .
Последним шагом в создании нашей молекулярной транспортной системы является высвобождение поршня из места его стыковки в центральном отверстии полностью собранной и закрытой гусеницы. Во время первых итераций проектирования нашей системы (дополнительный рис. 10) поршневой мономер имел очень низкую подвижность по нескольким причинам, поэтому было трудно судить, был ли выпуск успешным, путем отслеживания отдельных частиц в режиме реального времени.Поэтому мы прибегли к мониторингу вытекания поршней из стволов и гусениц с помощью ЭМА, отрицательного окрашивания ПЭМ и флуоресцентной микроскопии. То есть мы получили изображения нагруженных поршнем, но незакрытых дорожек до и после воздействия на образцы в течение нескольких часов или дней предполагаемых условий высвобождения поршня (дополнительный рисунок 23). Используя эту стратегию, мы определили процедуры, которые успешно вызывали высвобождение поршня на месте, даже несмотря на то, что фактическая диффузионная подвижность была слишком низкой, чтобы ее можно было увидеть в реальном времени в этот момент.В нашем окончательном дизайнерском решении мы освободили поршень от места его стыковки внутри закрытых гусениц, добавив пряди захвата снаружи. Нити захватчика проникают через межспиральные полости в стенках нити и освобождают поршень за счет смещения нити, опосредованного упором (дополнительная рис.
10) поршневой мономер имел очень низкую подвижность по нескольким причинам, поэтому было трудно судить, был ли выпуск успешным, путем отслеживания отдельных частиц в режиме реального времени.Поэтому мы прибегли к мониторингу вытекания поршней из стволов и гусениц с помощью ЭМА, отрицательного окрашивания ПЭМ и флуоресцентной микроскопии. То есть мы получили изображения нагруженных поршнем, но незакрытых дорожек до и после воздействия на образцы в течение нескольких часов или дней предполагаемых условий высвобождения поршня (дополнительный рисунок 23). Используя эту стратегию, мы определили процедуры, которые успешно вызывали высвобождение поршня на месте, даже несмотря на то, что фактическая диффузионная подвижность была слишком низкой, чтобы ее можно было увидеть в реальном времени в этот момент.В нашем окончательном дизайнерском решении мы освободили поршень от места его стыковки внутри закрытых гусениц, добавив пряди захвата снаружи. Нити захватчика проникают через межспиральные полости в стенках нити и освобождают поршень за счет смещения нити, опосредованного упором (дополнительная рис. 24). Большинство поршней в пределах поля зрения были подвижными в течение нескольких минут инкубации с нитями захватчика при комнатной температуре.
24). Большинство поршней в пределах поля зрения были подвижными в течение нескольких минут инкубации с нитями захватчика при комнатной температуре.
Наблюдение за движениями поршня в режиме реального времени
На подготовленных таким образом образцах мы наблюдали множество нитей с поршневыми узлами, совершающими случайные диффузионные движения вдоль дорожек, на которых они были ограничены (рис.3d, дополнительный фильм 1, дополнительные рис. 25–36). Количество мобильных единиц на гусенице можно контролировать с помощью начальной стехиометрии от поршневых бочек до бочек для полимеризации мультимеров. Следовательно, могут быть созданы ситуации, когда несколько поршней движутся по одной и той же дорожке, что приводит к ситуациям, когда они явно сталкиваются друг с другом, какое-то время движутся вместе, а затем расходятся (дополнительные фильмы 2 и 3).
Мы провели количественный анализ движения мобильных единиц по их следам, используя отслеживание центроидов со сверхвысоким разрешением 27 , что позволило определить положение во времени (рис. 4а, б, дополнительные рис. 25–36). Поршни имели несколько флуорофоров, которые позволяли непрерывно отслеживать частицы, как правило, в течение примерно ~ 10 минут, прежде чем сигнал стал слишком тусклым из-за обесцвечивания красителя.
4а, б, дополнительные рис. 25–36). Поршни имели несколько флуорофоров, которые позволяли непрерывно отслеживать частицы, как правило, в течение примерно ~ 10 минут, прежде чем сигнал стал слишком тусклым из-за обесцвечивания красителя.
a , b Типовые следы одиночных частиц вдоль самого дальнего измеренного расстояния (3 мкм) и с самой высокой измеренной подвижностью соответственно. c Энергетические профили, рассчитанные на основе распределений вероятности положения для трасс в a , b . d Красная пунктирная линия: приложенное извне напряжение, создающее электрическое поле вдоль нити накала. Сплошная линия: образцовый одночастичный след поршня, приводимого в движение полем.
Наблюдались диффузионные движения одиночных поршней по всей длине нижележащих нитей. Самые дальние зарегистрированные нами движения происходили на общей длине 3 мкм (рис.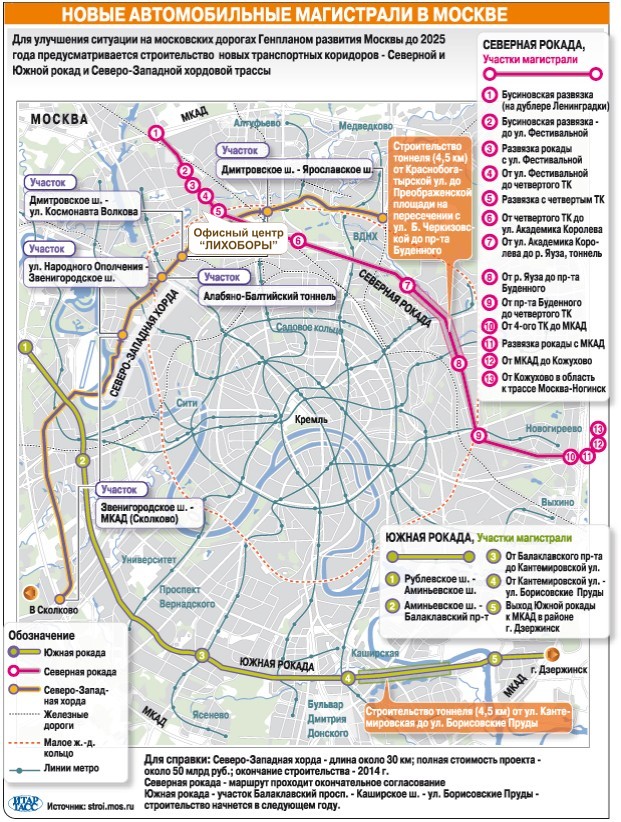 4а, дополнительный фильм 4). Отмечалась неоднородность по диффузионной подвижности частиц. Некоторые частицы будут неоднократно застревать в законсервированных местах (например,ж., рис. 4а), которые мы связываем с локальными блокпостами на путях. Такие дефекты могли быть вызваны одноцепочечными участками или слегка угловатыми соединениями между мономерами ствола, что потребовало бы энергетически невыгодной изгибной деформации поршня, чтобы пройти через такое сужение.
4а, дополнительный фильм 4). Отмечалась неоднородность по диффузионной подвижности частиц. Некоторые частицы будут неоднократно застревать в законсервированных местах (например,ж., рис. 4а), которые мы связываем с локальными блокпостами на путях. Такие дефекты могли быть вызваны одноцепочечными участками или слегка угловатыми соединениями между мономерами ствола, что потребовало бы энергетически невыгодной изгибной деформации поршня, чтобы пройти через такое сужение.
Другие частицы действительно демонстрировали очень высокую подвижность, перемещаясь на микрометровые расстояния за доли секунд, не застревая (рис. 4b, дополнительный фильм 5). Мы использовали позиционно-временные следы одной частицы, чтобы вычислить плотность вероятности для заполнения определенных положений нитей, а на основе этих данных с помощью Больцмана-инверсии профили свободной энергии следов (рис.4с). Профили свободной энергии иллюстрируют локальные минимумы и барьеры, на которых частицы, по-видимому, неоднократно захватывались.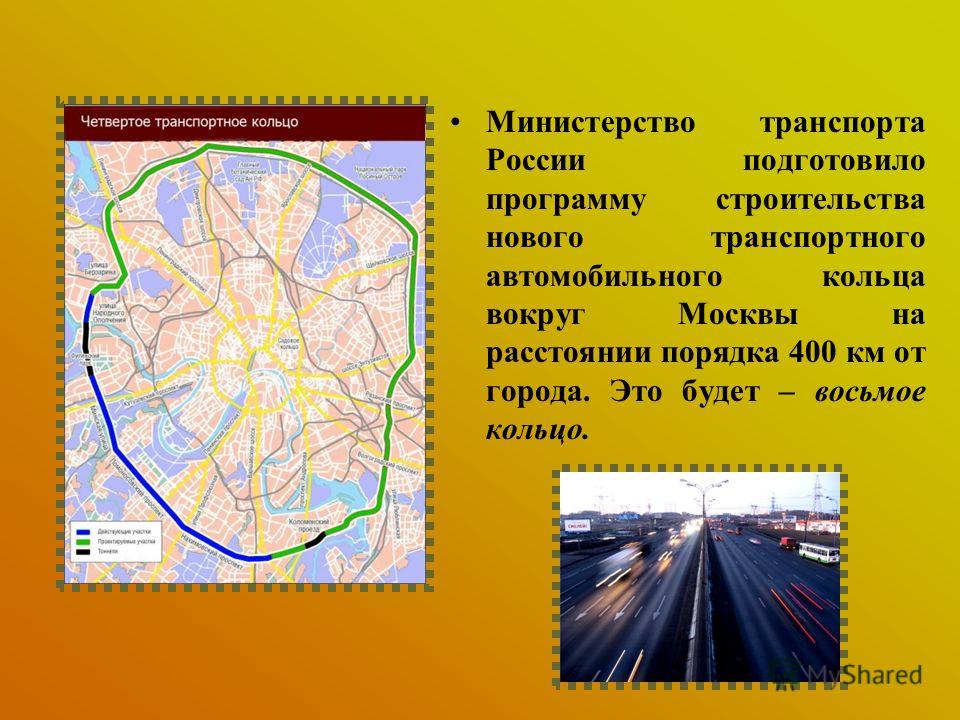 Например, высокомобильная частица на рис. 4b столкнулась с несколькими барьерами, причем самый высокий из них имел высоту всего ~3 k B T. Напротив, частица на рис. 4а, которая неоднократно оставалась застрявшей, имела на своем пути больше блокпостов с барьерами высотой до 5 k B T.
Например, высокомобильная частица на рис. 4b столкнулась с несколькими барьерами, причем самый высокий из них имел высоту всего ~3 k B T. Напротив, частица на рис. 4а, которая неоднократно оставалась застрявшей, имела на своем пути больше блокпостов с барьерами высотой до 5 k B T.
Управление однонаправленным движением поршня
Чтобы увидеть, могут ли внешние силы управлять движением поршня и потенциально ускорять преодоление препятствий, мы также отслеживали движение частиц в присутствии приложенных электрических полей.Два электрода в ранее описанной установке 28 использовались для создания электрического поля, направление которого менялось на противоположное каждые 5 с (рис. 4d). Мы заметили, что частицы на треках, которые были ориентированы параллельно полю, быстро двигались в ответ на поле (дополнительный рис. 37, дополнительные фильмы 6 и 7). Это означает, что при переключении поля поршни быстро перемещались от одного конца нижележащей нити к другому и затем оставались там до тех пор, пока поле не переключалось в противоположном направлении. При выключении поля поршни демонстрировали поведение типа диффузии с ловушками. Эти данные свидетельствуют о том, что захват вызван постоянными особенностями дорожки и не может быть устранен простым силовым вытягиванием поршня вдоль всей дорожки. Для поршней в гусеницах, которые были ориентированы перпендикулярно полю, как и ожидалось, поле практически не влияло на движение (дополнительный рисунок 38).
При выключении поля поршни демонстрировали поведение типа диффузии с ловушками. Эти данные свидетельствуют о том, что захват вызван постоянными особенностями дорожки и не может быть устранен простым силовым вытягиванием поршня вдоль всей дорожки. Для поршней в гусеницах, которые были ориентированы перпендикулярно полю, как и ожидалось, поле практически не влияло на движение (дополнительный рисунок 38).
Мобильность и лежащий в основе энергетический ландшафт
Чтобы количественно определить эффективную (свободную) диффузионную подвижность, мы вычислили среднеквадратичное смещение (СКО) за временные интервалы по временным трассам положения одной частицы (рис.5а–в). Кривые МСД сначала линейны во времени, как и ожидалось для нормальной диффузии, но затем они насыщаются. Насыщение отражает то, что диффузия происходит в треках конечного размера. Для нашей системы длина удержания соответствует всей длине нити, т. е. расстоянию между основными блокпостами. Анализируя множество частиц, мы обнаружили, что большинство частиц имеет коэффициент диффузии до 0,1 мкм 2 /с, но самые быстрые зарегистрированные частицы, у которых практически не было видимых препятствий, двигались со скоростью до 0.![]() 3 мкм 2 /с (рис. 5б). Диффузионная подвижность увеличивалась с повышением температуры окружающей среды, как видно из сравнения СКО от одиночных частиц, зарегистрированных при 20°, 25°, 30° и 35 °C (рис. 5c). Коэффициент диффузии примерно удваивается при переходе от 20 °C к 35 °C.
3 мкм 2 /с (рис. 5б). Диффузионная подвижность увеличивалась с повышением температуры окружающей среды, как видно из сравнения СКО от одиночных частиц, зарегистрированных при 20°, 25°, 30° и 35 °C (рис. 5c). Коэффициент диффузии примерно удваивается при переходе от 20 °C к 35 °C.
a Кривые среднего квадратического смещения (MSD) следов одиночных частиц на рис. 4a, b соответственно (рассчитано в соответствии с дополнительным файлом программного обеспечения 3). b Гистограмма коэффициентов диффузии D N = 128 частиц. c Символы: Среднее СКО как функция временных интервалов движения одиночной частицы, зарегистрированное при указанных температурах. 20 ° C: N = 28, 25 ° C: N = 32, 30 ° C: N = 37, 35 ° C: N = 31. D сумма пространственного автокорреляции вычисляется на н. = 128 частиц. e Круги: распределения скоростей, вычисленные на основе производной по времени следов положения одной частицы во времени. Пунктирная линия: распределение Гаусса. Красная сплошная линия: распределение скоростей, смоделированное с использованием динамики Ланжевена (рассчитано в соответствии с дополнительными программными файлами 1 и 2). Врезка: поверхность свободной энергии, используемая в моделировании.
Пунктирная линия: распределение Гаусса. Красная сплошная линия: распределение скоростей, смоделированное с использованием динамики Ланжевена (рассчитано в соответствии с дополнительными программными файлами 1 и 2). Врезка: поверхность свободной энергии, используемая в моделировании.
Мы рассчитали пространственную автокорреляцию плотности вероятности, чтобы заполнить позиции треков от каждой зарегистрированной частицы, чтобы исследовать скрытые периодичности. Пространственная автокорреляционная функция (рис. 5d), усредненная по многим записям отдельных частиц, показывает четкие периодические пики, возникающие в интервалах 64 нм, что соответствует длине одной бочкообразной субъединицы.Периодичность можно увидеть и в пространственной автокорреляции от отдельных частиц, хотя и менее отчетливо. Тот факт, что расчетная периодичность трека восстанавливается из движения частиц, предполагает, что подвижные единицы на мгновение попадают в ловушку периодически возникающих структурных особенностей. Такое поведение может быть вызвано, например, местом стыковки поршня (небольшое углубление), которое появляется в каждом цилиндрическом мономере.
Такое поведение может быть вызвано, например, местом стыковки поршня (небольшое углубление), которое появляется в каждом цилиндрическом мономере.
Отличительной чертой броуновского движения является гауссово распределение скоростей.Напротив, распределение скоростей, вычисленное по экспериментальным траекториям положения и времени случайного блуждающего устройства, сильно отклоняется от гауссова (рис. 5e). Негауссовское поведение распределений, усредненных по отдельным частицам, не обязательно означает, что движение одиночной свободно диффундирующей частицы следует негауссовскому процессу, а может быть результатом отклонений среди отдельных частиц 29 . Вместо этого в данном случае мы подозреваем, что дополнительные негауссовские эффекты исходят из периодического потенциального ландшафта, связанного с молекулярной структурой ствола.Чтобы еще больше обосновать это, мы сравниваем наши экспериментальные результаты с ланжевеновским моделированием частицы в квазипериодическом потенциале (рис. 5e, вставка).
5e, вставка).
В имитационной модели частица рассеивается под действием случайного белого шума, имитирующего тепловую стохастическую среду. Стандартное отклонение скорости частиц сильно зависит от высоты барьера (см. Дополнительное примечание 1). Анализ экспериментальных следов свидетельствует о значительном варьировании потенциальных барьеров (т.г., рис. 4а и 4б). Соответственно, мы усреднили смоделированные траектории с потенциальными высотами барьеров, которые варьируются в диапазоне от 1 до 10 k B T. Чтобы соответствовать экспериментальным условиям, мы также добавили локализационный шум к положениям смоделированных траекторий. При фиксированном периоде a = 64 нм и фиксированной ширине потенциальной ямы b = 4 нм распределение скоростей, полученное усреднением по многим смоделированным таким образом одночастичным траекториям, хорошо согласуется с экспериментально наблюдаемой скоростью распределение, которое показывает, что негауссово поведение связано с периодическими потенциальными ландшафтами с различной высотой барьера.
Чем отличается транспортировка и хранение воды в ранней и поздней древесине хвойных пород? | Журнал экспериментальной ботаники
Аннотация
Цель этого исследовательского проекта состояла в том, чтобы определить поведение водного транспорта ранней древесины по сравнению с поздней в стволе 21-летней пихты Дугласа [ Pseudostuga menziesii (Mirb.) Franco]. Удельная электропроводность ( k с ) и уязвимость ксилемы к эмболии были измерены на одном годичном кольце и в подмножестве образцов ранней и поздней древесины в одном кольце.Соотношение ранней/поздней древесины, водный потенциал ствола (Ψ) и относительное содержание воды ( RWC ) использовались для прогнозирования различий в проводимости и уязвимости к эмболии. В ранней древесине примерно в 11 раз больше k s , чем в поздней древесине, и до 90% общего потока приходится на раннюю древесину. Уязвимость ранней древесины к эмболии соответствовала той же тенденции, что и у всей древесины, с 50% потерей проводимости при -2,2 МПа ( P 50 ). Поздняя древесина была более уязвима для эмболии, чем ранняя древесина при высоком Ψ, но по мере уменьшения Ψ поздняя древесина демонстрировала очень небольшую дальнейшую эмболию, с P 50 <–5.0 МПа. Самый низкий Ψ ствола, оцененный в полевых условиях, составлял около –1,4 МПа, что указывает на то, что поздняя и ранняя древесина в полевых условиях потеряли около 42% и 16% соответственно. Более высокая уязвимость к эмболии в поздней древесине, чем в ранней древесине на поле Ψ, была связана с более высокой водоаккумулирующей способностью (21,8% RWC МПа –1 против 4,1% RWC МПа –1 , поздняя и ранняя древесина соответственно). Форма кривой уязвимости свидетельствует о том, что засев воздуха через позднюю древесину может происходить непосредственно через поры в марго и закрываться при более низком давлении, чем поры ранней древесины.
Поздняя древесина была более уязвима для эмболии, чем ранняя древесина при высоком Ψ, но по мере уменьшения Ψ поздняя древесина демонстрировала очень небольшую дальнейшую эмболию, с P 50 <–5.0 МПа. Самый низкий Ψ ствола, оцененный в полевых условиях, составлял около –1,4 МПа, что указывает на то, что поздняя и ранняя древесина в полевых условиях потеряли около 42% и 16% соответственно. Более высокая уязвимость к эмболии в поздней древесине, чем в ранней древесине на поле Ψ, была связана с более высокой водоаккумулирующей способностью (21,8% RWC МПа –1 против 4,1% RWC МПа –1 , поздняя и ранняя древесина соответственно). Форма кривой уязвимости свидетельствует о том, что засев воздуха через позднюю древесину может происходить непосредственно через поры в марго и закрываться при более низком давлении, чем поры ранней древесины.
Поступила 10 мая 2002 г.; Принято 23 июля 2002 г.
Введение
В годичном кольце есть две отдельные зоны роста: ранняя древесина и поздняя древесина.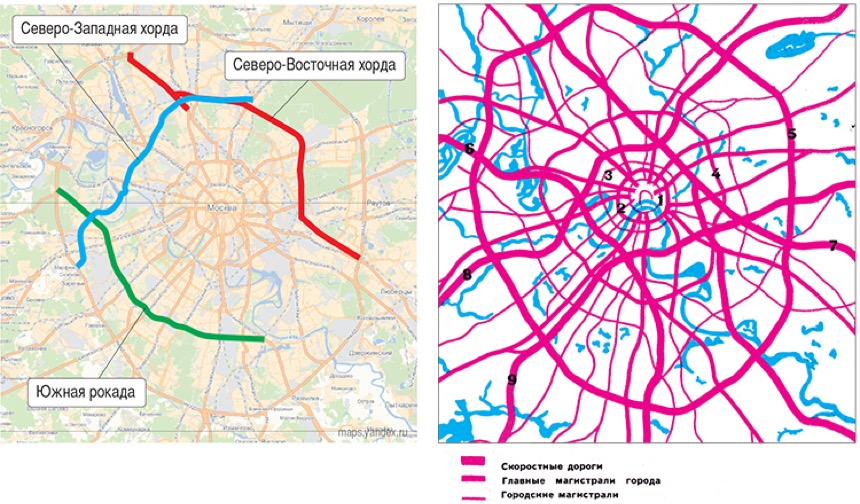 На раннесезонный прирост дерева, образующего раннюю древесину (периклинальный рост), приходится 40–80 % прироста кольца в ширину за год. Поскольку скорость роста замедляется и останавливается осенью, отложенные клетки становятся меньше, плотнее и механически прочнее (поэтому кажутся темнее).
На раннесезонный прирост дерева, образующего раннюю древесину (периклинальный рост), приходится 40–80 % прироста кольца в ширину за год. Поскольку скорость роста замедляется и останавливается осенью, отложенные клетки становятся меньше, плотнее и механически прочнее (поэтому кажутся темнее).
Вода является наиболее ограниченным экологическим ресурсом для большинства деревьев и лесных массивов (Richter, 1976; Waring and Running, 1998).Изменения в характере роста ранней/поздней древесины отчасти отражают контроль свойств древесины за счет наличия воды. Такие факторы, как температура, условия солнечного освещения и уклон склона холма, также могут влиять на ширину годичных колец и количество ранней или поздней древесины (Larson, 1962). Многочисленные исследования показали, что у хвойных в целом более высокая плотность древесины вследствие большей доли поздней древесины вызывается стрессом от засухи (см. обзор Zobel and Van Buijtenen, 1989).Поздняя древесина более чувствительна к климату, чем ранняя (Bannan, 1967; Lebourgeois, 2000). В вегетационные периоды с меньшим количеством осадков, чем обычно, и чрезмерно низкими температурами рост деревьев уменьшается, а доля поздней древесины выше. И наоборот, когда условия выращивания идеальны, рост деревьев усиливается, а доля поздней древесины уменьшается (Кеннеди, 1961).
В вегетационные периоды с меньшим количеством осадков, чем обычно, и чрезмерно низкими температурами рост деревьев уменьшается, а доля поздней древесины выше. И наоборот, когда условия выращивания идеальны, рост деревьев усиливается, а доля поздней древесины уменьшается (Кеннеди, 1961).
Хотя значение наружного кольца для гидравлической проводимости, а ранней и поздней древесины для ксилемной эмболии всего ствола было количественно определено для кольцевидных пористых деревьев ( Ulmus americana , Ellmore and Ewers, 1986; Quercus cerris, Lo Gullo и др. ., 1995), ни одно исследование не исследовало его у хвойных пород или в ранней и поздней древесине одного и того же кольца. Кроме того, нет сообщений о прямом измерении уязвимости к эмболии в пределах одного кольца и его компонентов. Логично предположить, что ранняя и поздняя древесина будут иметь очень разные гидравлические свойства.
Различия в гидравлической проводимости и подверженности эмболии между ранней и поздней древесиной связаны с различиями в анатомии ксилемы (Zimmermann, 1983). Гидравлическая проводимость пропорциональна четвертой степени радиуса каналов ксилемы (Gibson и др. , 1985; Tyree and Ewers, 1991). Трахеиды ранней древесины имеют более широкий диаметр просвета, чем трахеиды поздней древесины (Panshin and de Zeuuw, 1980; Ewers et al. ., 1997). Компромисс, однако, заключается в том, что каналы с высокой проводимостью могут быть более уязвимыми для эмболии (Schulte and Gibson, 1988; Sperry and Saliendra, 1994; Hargrave et al ., 1994). Эти данные свидетельствуют о том, что ранняя древесина может переносить большую часть воды к основному стволу при благоприятном водном статусе растений, но может ограничивать использование воды растениями во время засухи.Считается, что в лиственных породах сосуды поздней древесины малого диаметра и сосудоцентрические трахеиды малого диаметра рядом с более крупными сосудами обеспечивают транспорт воды после того, как ранняя древесина и/или более широкие сосуды наполняются воздухом (Carlquist, 1985; Cochard et al .
Гидравлическая проводимость пропорциональна четвертой степени радиуса каналов ксилемы (Gibson и др. , 1985; Tyree and Ewers, 1991). Трахеиды ранней древесины имеют более широкий диаметр просвета, чем трахеиды поздней древесины (Panshin and de Zeuuw, 1980; Ewers et al. ., 1997). Компромисс, однако, заключается в том, что каналы с высокой проводимостью могут быть более уязвимыми для эмболии (Schulte and Gibson, 1988; Sperry and Saliendra, 1994; Hargrave et al ., 1994). Эти данные свидетельствуют о том, что ранняя древесина может переносить большую часть воды к основному стволу при благоприятном водном статусе растений, но может ограничивать использование воды растениями во время засухи.Считается, что в лиственных породах сосуды поздней древесины малого диаметра и сосудоцентрические трахеиды малого диаметра рядом с более крупными сосудами обеспечивают транспорт воды после того, как ранняя древесина и/или более широкие сосуды наполняются воздухом (Carlquist, 1985; Cochard et al . , 1997). В хвойных породах вопрос остается нерешенным. Остается ли поздняя древесина проводящей после того, как ранняя древесина наполняется воздухом, или нет?
, 1997). В хвойных породах вопрос остается нерешенным. Остается ли поздняя древесина проводящей после того, как ранняя древесина наполняется воздухом, или нет?
Эмболия из-за засухи, как полагают, вызвана засевом воздуха в мембранах межтрубных ямок (Zimmermann, 1983; Crombie et al ., 1985). Засев воздуха у хвойных происходит через мембрану межтрахеидной ямки, когда тор смещается из своего нормального положения уплотнения (Sperry and Tyree, 1990). Они обнаружили, что незапечатывание окаймленных ямок в поздней древесине у хвойных может составлять 20% общей гидравлической проводимости после того, как ранняя древесина запечаталась при низком водном потенциале (Ψ). Форма кривых уязвимости различается между видами и внутри них, и считается, что они отражают среду водного стресса, в которой росли растения (Тайри и Сперри, 1988; Тайри и др. )., 1994; Matzner и др. , 2001).
Гораздо меньше известно о реакции ранней и поздней древесины на эмболию, а также о степени эмболии, возникающей в этих двух зонах кольца роста в поле Ψ.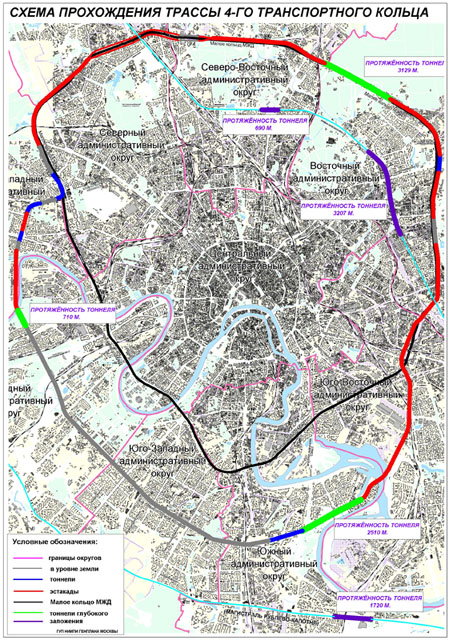 Соотношение ранней и поздней древесины в пределах кольца и анатомии ксилемы может быть адаптивным гидравлическим свойством, а не просто следствием водного баланса в поле. Ранняя и поздняя древесина могут регулировать перенос воды в условиях стресса, основываясь на их влиянии на гидравлическую проводимость, уязвимость к эмболии и внутреннюю накопительную способность.Предполагается, что на анатомической основе, как и предсказывает гипотеза «засева воздуха», гидравлические функции напрямую связаны с функционированием окаймленных ямок и механическими свойствами структуры мембраны ямок, предложенной Петти (1972). Эмболия, развивающаяся в трахеиде ранней древесины, не будет распространяться на соседние трахеиды, потому что давление воздуха, создаваемое между заполненной водой трахеидой и пустой трахеидой, будет превышать механическое напряжение, необходимое для протягивания пузырька воздуха через марго.Тор запечатывает камеру до того, как воздух сможет пройти через поры. Следовательно, снижение гидравлической проводимости в ранней древесине будет связано с механическими свойствами мембраны.
Соотношение ранней и поздней древесины в пределах кольца и анатомии ксилемы может быть адаптивным гидравлическим свойством, а не просто следствием водного баланса в поле. Ранняя и поздняя древесина могут регулировать перенос воды в условиях стресса, основываясь на их влиянии на гидравлическую проводимость, уязвимость к эмболии и внутреннюю накопительную способность.Предполагается, что на анатомической основе, как и предсказывает гипотеза «засева воздуха», гидравлические функции напрямую связаны с функционированием окаймленных ямок и механическими свойствами структуры мембраны ямок, предложенной Петти (1972). Эмболия, развивающаяся в трахеиде ранней древесины, не будет распространяться на соседние трахеиды, потому что давление воздуха, создаваемое между заполненной водой трахеидой и пустой трахеидой, будет превышать механическое напряжение, необходимое для протягивания пузырька воздуха через марго.Тор запечатывает камеру до того, как воздух сможет пройти через поры. Следовательно, снижение гидравлической проводимости в ранней древесине будет связано с механическими свойствами мембраны.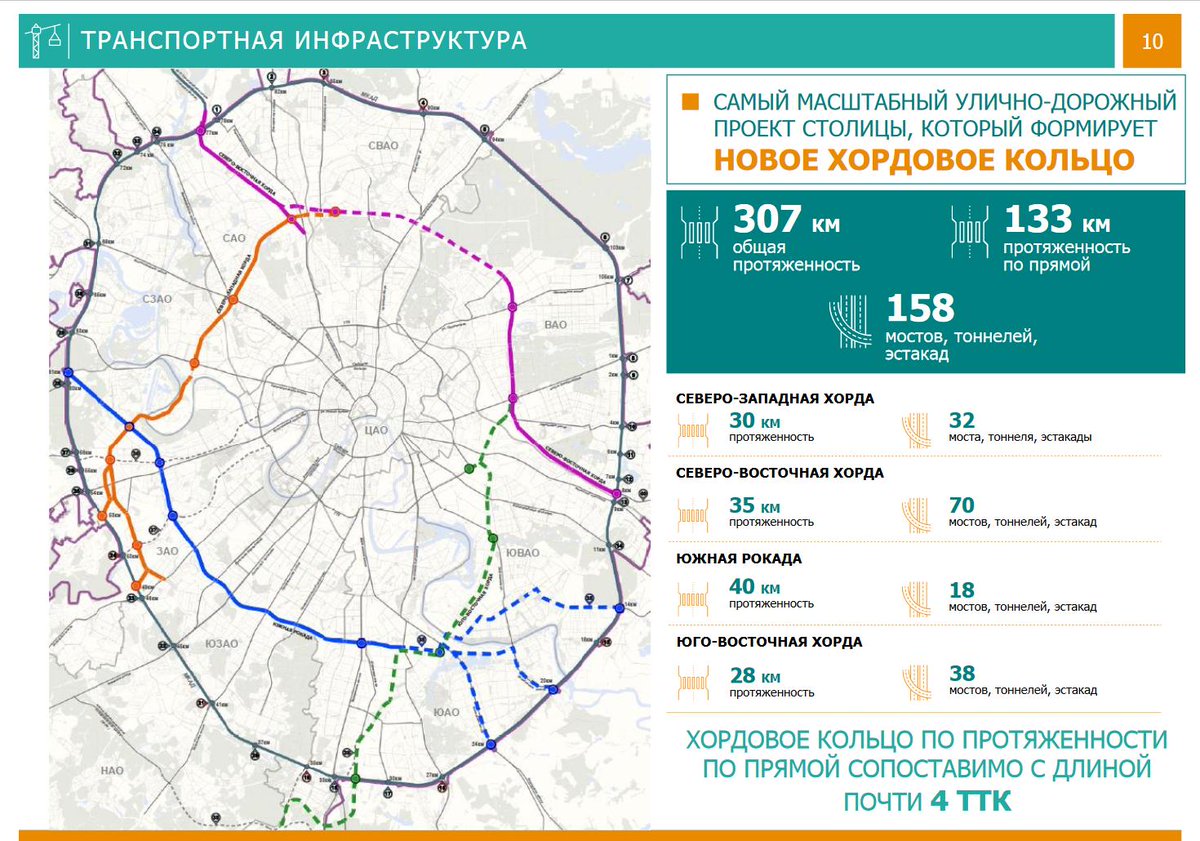 В поздней древесине воздух будет проходить через поры до того, как тор можно будет запечатать из-за его высокой механической устойчивости к смещению. Таким образом, снижение гидравлической проводимости поздней древесины будет в большей степени зависеть от размера пор, проходящих через марго. Поэтому ожидается, что из-за их окаймленной ямочной структуры трахеиды ранней древесины будут более эффективными и более безопасными от эмболии, чем поздняя древесина.Если, как и ожидалось, устойчивость ранней древесины к эмболии выше, безопасная ксилема не требует больших затрат с точки зрения инвестиций в углерод.
В поздней древесине воздух будет проходить через поры до того, как тор можно будет запечатать из-за его высокой механической устойчивости к смещению. Таким образом, снижение гидравлической проводимости поздней древесины будет в большей степени зависеть от размера пор, проходящих через марго. Поэтому ожидается, что из-за их окаймленной ямочной структуры трахеиды ранней древесины будут более эффективными и более безопасными от эмболии, чем поздняя древесина.Если, как и ожидалось, устойчивость ранней древесины к эмболии выше, безопасная ксилема не требует больших затрат с точки зрения инвестиций в углерод.
Целью данного исследования было сравнение способности ранней и поздней древесины транспортировать воду в пределах одного кольца роста у быстрорастущих деревьев дугласовой пихты [ Pseudostuga menziesii (Mirb.) Franco]. Гидравлические параметры (электропроводность, уязвимость к эмболии, водоаккумулирующая способность) измеряли на целых образцах древесины, а также на образцах ранней и поздней древесины. Полевые данные измерений Ψ и относительного содержания воды использовались для подтверждения лабораторных результатов и для описания транспортировки и накопления воды в ранней и поздней древесине, когда вода находится под давлением.
Полевые данные измерений Ψ и относительного содержания воды использовались для подтверждения лабораторных результатов и для описания транспортировки и накопления воды в ранней и поздней древесине, когда вода находится под давлением.
Это исследование было разработано для проверки следующих трех гипотез: (1) электропроводность заболони в поздней и цельной древесине ниже, чем в ранней древесине, (2) электропроводность ранней и поздней древесины, взвешенная по соотношению между ранней и поздней древесиной, объясняет электропроводность всей древесины. и (3) подверженность эмболии выше у поздней древесины, чем у ранней.
Материалы и методы
Растительный материал и подготовка проб
Исследование проводилось в одновозрастном (21 год) насаждении дугласовой пихты (высота 277 м, 44°43′ с.ш., 123°20′ з.д.) в 15 км к северо-западу от Корваллиса, штат Орегон, растущего на инцептизольная почва. Среднегодовая температура и годовое количество осадков (1961–1996 гг.) составляют 12,4 °C и 1800 мм соответственно (таблица 1; www.orst.edu/Dept/IPPC/wea/). Дугласова пихта была выбрана из-за ее быстрого роста, что дает широкие годичные кольца с ярко выраженной поздней древесиной.В середине июня 2001 г. (до начала летней засухи) случайным образом были отобраны и срублены пять деревьев. От каждого дерева вырезали диск толщиной 20 см на уровне груди и сразу же транспортировали во влажном полиэтиленовом пакете в лабораторию, где готовили блоки. Из каждого диска в поперечном сечении шесть блоков с интервалами примерно 60° по окружности были вырезаны в одном и том же кольце с помощью молотка и долота (Domec and Gartner, 2001).
Среднегодовая температура и годовое количество осадков (1961–1996 гг.) составляют 12,4 °C и 1800 мм соответственно (таблица 1; www.orst.edu/Dept/IPPC/wea/). Дугласова пихта была выбрана из-за ее быстрого роста, что дает широкие годичные кольца с ярко выраженной поздней древесиной.В середине июня 2001 г. (до начала летней засухи) случайным образом были отобраны и срублены пять деревьев. От каждого дерева вырезали диск толщиной 20 см на уровне груди и сразу же транспортировали во влажном полиэтиленовом пакете в лабораторию, где готовили блоки. Из каждого диска в поперечном сечении шесть блоков с интервалами примерно 60° по окружности были вырезаны в одном и том же кольце с помощью молотка и долота (Domec and Gartner, 2001).
Образцы вырезали из древесины так, чтобы длинная ось была параллельна волокнам, затем погружали в воду и помещали в вакуум на 48 часов, чтобы заполнить часть эмболизированных трахеид.Их первоначальные размеры составляли 150 мм (в осевом направлении), 10 мм в тангенциальном направлении и 7–9 мм в радиальном направлении (ширина одного кольца роста). Образцы были случайным образом обозначены как A–F. Все гидравлические измерения образца А проводились на всем образце, т. е. как ранняя, так и поздняя древесина оставались нетронутыми. Перед окончательными измерениями образцы B–F были распилены пополам до 75 мм в осевом направлении. Одна половина была обозначена B 1 –F 1 , а другая половина B 2 –F 2 .С помощью долота и бритвенных лезвий из образцов В 1 – F 1 удалили позднюю древесину, оставив только раннюю древесину, а раннюю древесину удалили из образцов В 2 – F 2 , оставив только позднюю древесину (рис. 1).
Образцы были случайным образом обозначены как A–F. Все гидравлические измерения образца А проводились на всем образце, т. е. как ранняя, так и поздняя древесина оставались нетронутыми. Перед окончательными измерениями образцы B–F были распилены пополам до 75 мм в осевом направлении. Одна половина была обозначена B 1 –F 1 , а другая половина B 2 –F 2 .С помощью долота и бритвенных лезвий из образцов В 1 – F 1 удалили позднюю древесину, оставив только раннюю древесину, а раннюю древесину удалили из образцов В 2 – F 2 , оставив только позднюю древесину (рис. 1).
Удельная и гидравлическая проводимость
Удельная проводимость ( k с, в м 2 ) является мерой гидравлической эффективности ксилемы по отношению к площади поперечного сечения древесины и рассчитывается в соответствии с законом Дарси (Эдвардс и Джарвис).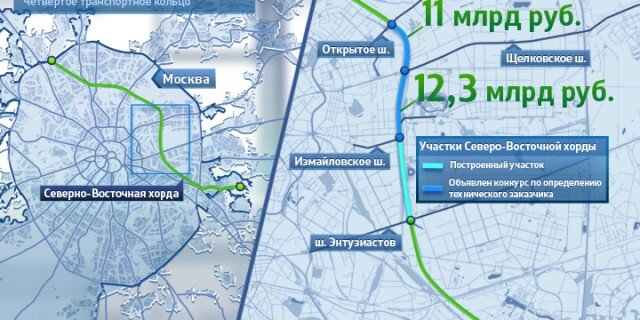 , 1982).Начальная удельная электропроводность ( k с(i) ) была измерена на каждом сегменте 130–150 мм (неповрежденные сегменты A–F, до того, как они были распилены пополам) с использованием мембранного напорного рукава, необходимого для обеспечения того, чтобы жидкость не утечка по бокам образцов (Spicer and Gartner, 1998 a ). Образцы замачивали в воде, затем измеряли k s(i) при перепаде давления 2,8 кПа с использованием фильтрованной (0,22 мкм) воды, доведенной с помощью HCl до pH 2 для предотвращения микробного роста.Гидравлический напор был предварительно выбран достаточно низким, чтобы избежать повторного наполнения эмболизированных трахеид. Эфлюкс собирали микропипеткой с градуировкой на 1 мл (градуировка 0,01 мл). Когда поток стабилизировался, регистрировали время, необходимое для пересечения мениском шести последовательных делений деления. Гидравлическая проводимость, k ч (в м 4 ), которая выражает отношение объемного расхода к давлению, но не на основе площади (Tyree and Ewers, 1991), была определена как k ч = A × k s , где A — площадь поперечного сечения (м 2 ) образца.
, 1982).Начальная удельная электропроводность ( k с(i) ) была измерена на каждом сегменте 130–150 мм (неповрежденные сегменты A–F, до того, как они были распилены пополам) с использованием мембранного напорного рукава, необходимого для обеспечения того, чтобы жидкость не утечка по бокам образцов (Spicer and Gartner, 1998 a ). Образцы замачивали в воде, затем измеряли k s(i) при перепаде давления 2,8 кПа с использованием фильтрованной (0,22 мкм) воды, доведенной с помощью HCl до pH 2 для предотвращения микробного роста.Гидравлический напор был предварительно выбран достаточно низким, чтобы избежать повторного наполнения эмболизированных трахеид. Эфлюкс собирали микропипеткой с градуировкой на 1 мл (градуировка 0,01 мл). Когда поток стабилизировался, регистрировали время, необходимое для пересечения мениском шести последовательных делений деления. Гидравлическая проводимость, k ч (в м 4 ), которая выражает отношение объемного расхода к давлению, но не на основе площади (Tyree and Ewers, 1991), была определена как k ч = A × k s , где A — площадь поперечного сечения (м 2 ) образца.
Кривые уязвимости и емкость для хранения воды
Кривые уязвимости (VC) образцов древесины были построены с помощью метода нагнетания воздуха (Salleo et al. ., 1992; Sperry and Saliendra, 1994), адаптированного для образцов, взятых непосредственно из ствола (Domec and Gartner, 2001) . Метод нагнетания воздуха основан на теории, согласно которой отрицательный водный потенциал (Ψ), необходимый для втягивания воздуха в канал и вызывающий эмболию, равен положительному давлению воздуха, необходимому для проталкивания воздуха в канал, когда Ψ равно атмосферному давлению.Цель состояла в том, чтобы измерить потерю проводимости в процентах ( PLC ) на сегментах путем попеременного перемещения образца между напорным рукавом с мембраной для определения k s и двусторонней камерой давления для вызывания эмболии. Сегменты были вставлены в камеру для нагнетания воздуха, причем оба конца выступали наружу. Воздухонепроницаемая камера была герметизирована резиновыми прокладками с обоих концов, удерживаемыми на месте торцевыми крышками из нержавеющей стали. Давление в камере создавали ступенчато и поддерживали постоянным в течение 1 мин.После каждого нагнетания воздуха k s повторно измеряли, и процентное соотношение k s(i) было ниже k s(i) , что давало образец PLC . Температуру раствора, свежую массу ( M f ), объем ( V f ), определяемый вытеснением воды (принцип Архимеда), и длину каждой пробы регистрировали до и после каждой пробы. измерение проводимости. Образцы всегда имели одинаковый вес до и после, что не свидетельствовало о повторном наполнении. VC были определены путем построения графика зависимости PLC от Ψ, где Ψ прогнозируется как отрицательное значение приложенного давления воздуха (Pockman and Sperry, 2000).
Давление в камере создавали ступенчато и поддерживали постоянным в течение 1 мин.После каждого нагнетания воздуха k s повторно измеряли, и процентное соотношение k s(i) было ниже k s(i) , что давало образец PLC . Температуру раствора, свежую массу ( M f ), объем ( V f ), определяемый вытеснением воды (принцип Архимеда), и длину каждой пробы регистрировали до и после каждой пробы. измерение проводимости. Образцы всегда имели одинаковый вес до и после, что не свидетельствовало о повторном наполнении. VC были определены путем построения графика зависимости PLC от Ψ, где Ψ прогнозируется как отрицательное значение приложенного давления воздуха (Pockman and Sperry, 2000).
После измерения k s(i) на неповрежденных образцах A–F было построено VC s двумя способами. Во-первых, образец A был использован для изготовления всего VC , подвергая его воздействию давления воздуха в диапазоне от 0,5 до 5,0 МПа. Во-вторых, для построения одной кривой использовались разные образцы B–F при разном давлении воздуха (рис.2). После определения k s для образцов B и C их распилили пополам для получения образцов B 1 и C 1 (только ранняя древесина) и B 2 и C 2 (только поздняя древесина). ), и k s измеряли повторно (образцы B, C, B 1, B 2 , C 1, и C 2 ) использовали в качестве повторов для 5 8
Во-вторых, для построения одной кривой использовались разные образцы B–F при разном давлении воздуха (рис.2). После определения k s для образцов B и C их распилили пополам для получения образцов B 1 и C 1 (только ранняя древесина) и B 2 и C 2 (только поздняя древесина). ), и k s измеряли повторно (образцы B, C, B 1, B 2 , C 1, и C 2 ) использовали в качестве повторов для 5 8 k
при полном насыщении). После определения k s для образца D давление в нем было равно 0.5 МПа, его к с перемеряли, затем распиливали пополам для изготовления образцов Д 1 и Д 2 , на которых измеряли к с . Аналогичным образом, после определения k s для образцов E и F они были опрессованы до 1,0 и 3,0 МПа соответственно, повторно измерены их k s , затем они были распилены пополам на подготовленные образцы E ..jpg) 1 , E 2 , F 1 и F 2 , на которых было измерено k s .Для образцов ранней и поздней древесины PLC определяли методом парных сегментов, в котором сравнивали k s(i) образцов D 1 , D 2 , E 1 , E . , F 1 , F 2 к среднему k s(i) образцов B 1 , B 2 , C 1 1 из того же диска, (рис. 2). Чтобы проверить различия между методом парных сегментов и обычным методом (состоящим из измерения PLC в одних и тех же образцах при разных Ψ), было проведено сравнение при каждом Ψ между образцами A и интактными образцами B–F.
1 , E 2 , F 1 и F 2 , на которых было измерено k s .Для образцов ранней и поздней древесины PLC определяли методом парных сегментов, в котором сравнивали k s(i) образцов D 1 , D 2 , E 1 , E . , F 1 , F 2 к среднему k s(i) образцов B 1 , B 2 , C 1 1 из того же диска, (рис. 2). Чтобы проверить различия между методом парных сегментов и обычным методом (состоящим из измерения PLC в одних и тех же образцах при разных Ψ), было проведено сравнение при каждом Ψ между образцами A и интактными образцами B–F.
Площадь эмболии, оцененная методом окрашивания
После каждого окончательного измерения электропроводности каждый образец перфузировали отфильтрованным (0,22 мкм) сафранином-О (0,5% водный раствор) под напором 15 кПа в течение 20 минут, оставаясь в напорном рукаве.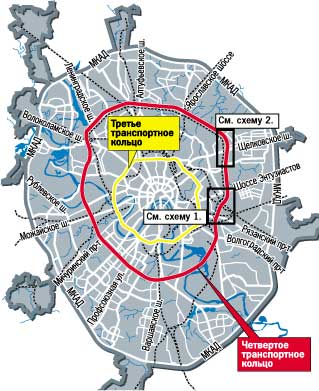 Гидравлический напор был заранее выбран достаточно высоким, чтобы позволить сафранину-О течь без повторного наполнения какой-либо из эмболизированных трахеид. Для первоначальных измерений образцы всегда полностью окрашивались, не давая признаков эмболии.Процент неокрашенной площади древесины каждого образца, использованного для VC s, был проанализирован с помощью системы анализа изображений, состоящей из составного микроскопа, видеокамеры, компьютера и программного обеспечения NIH Image (v. 1.60, Rasband, 1996). .
Гидравлический напор был заранее выбран достаточно высоким, чтобы позволить сафранину-О течь без повторного наполнения какой-либо из эмболизированных трахеид. Для первоначальных измерений образцы всегда полностью окрашивались, не давая признаков эмболии.Процент неокрашенной площади древесины каждого образца, использованного для VC s, был проанализирован с помощью системы анализа изображений, состоящей из составного микроскопа, видеокамеры, компьютера и программного обеспечения NIH Image (v. 1.60, Rasband, 1996). .
Доля ранней и поздней древесины
Суммарная ширина ранней древесины, поздней древесины и всего кольца определялась для каждого образца до того, как его половинки были вырублены на только позднюю или только раннюю древесину. Эти измерения были выполнены с помощью устройства для измерения годичных колец следующим образом.Образец помещали на подвижный измерительный стол Velmex, оснащенный линейным энкодером AcuRite ENC 150, цифровым измерительным устройством и дисплеем QuickCheck-1000. Наблюдая за изображением кольца роста на мониторе, проецирующем увеличенное видеоизображение образца, а затем перемещая предметный столик, можно определить ширину кольца роста с точностью до 0,01 мм. Затем рассчитывали среднюю долю ранней древесины для каждого образца.
Наблюдая за изображением кольца роста на мониторе, проецирующем увеличенное видеоизображение образца, а затем перемещая предметный столик, можно определить ширину кольца роста с точностью до 0,01 мм. Затем рассчитывали среднюю долю ранней древесины для каждого образца.
Содержание влаги, относительное содержание воды и запас воды
Содержание влаги ( MC ) определяется как масса воды, присутствующей в куске древесины, выраженная в процентах от массы высушенной в печи древесины ( M d ):
Относительное содержание воды ( RWC ) — это масса воды в образце, деленная на потенциально максимальные значения содержания воды в образце.Чтобы получить скорость изменения RWC , связанную с кавитацией, RWC определяли для каждого образца, использованного для создания VC , в начале и после каждого давления путем записи V f (см 3 ) и М ф (г). После окончательного повышения давления M d (g) после сушки при 104 °C. Длина использовалась для обратного расчета объема материала клеточной стенки ( V с , см 3 ) и M d (g) при каждом заданном давлении. RWC можно рассчитать следующим образом:
После окончательного повышения давления M d (g) после сушки при 104 °C. Длина использовалась для обратного расчета объема материала клеточной стенки ( V с , см 3 ) и M d (g) при каждом заданном давлении. RWC можно рассчитать следующим образом:
где D H 2 O – плотность воды (г см –3). V s был рассчитан из M d , предполагая, что материал стенок сухих ячеек поздней и ранней древесины имеет плотность 1,53 г см -3 (Kennedy and Warren, 1969; Kellog et al. 9018). , 1975; Сиау, 1984).
Влагоемкость, определяемая как количество воды, забираемой из ствола при заданном водном потенциале (Holbrook, 1995), выражалась как изменение RWC на единицу Ψ.Это определение емкости позволило сравнить различия в емкости хранения, которые связаны с различием не только в общем объеме воды, но и в плотности тканей (Domec, Gartner, 2001). Объемные емкости на основе RWC ( C RWC ) для заданного диапазона давлений определялись после каждой точки электропроводности следующим образом (Edwards and Jarvis, 1982): с использованием барокамеры, поскольку положительное давление не может вытеснить капиллярную воду (Zimmermann, 1983; Tyree and Yang, 1990).Гипотеза капиллярного накопления воды утверждает, что накопление воды происходит за счет изменения радиуса мениска между клеточной стенкой и газовым пространством (Zimmermann, 1983). Если эта гипотеза верна, то максимальное накопление капиллярной воды высвобождается при давлении, близком к нулю, и прекращается до 0,6 МПа (Holbrook, 1995; Tyree, Yang, 1990). Поэтому, чтобы определить, был ли наклон кривой дегидратации постоянным во всем ее диапазоне, и избежать недооценки капиллярной воды при низком приложенном давлении, значения емкости были рассчитаны для трех фаз на каждой кривой дегидратации: фаза от 0 до 0.5 МПа ( С 0–0,5 ), фаза от 0,5–1,0 МПа ( С 0,5–1,0 ) и фаза от 1,0–3,0 МПа ( С 09077 1,0–3,0 МПа).
Плотность древесины и объем, занимаемый материалом клеточных стенок, водой и газом
Значения плотности древесины были определены для каждого образца, испытанного гидравлически:
Для каждого типа образца объем, занимаемый водой, материалом клеточных стенок и газом (на основе свежего объема), был рассчитан как:
V GAS (%) = (1- V Стена сотовой связи + V H 2 O ) 100 (7)
2 Полевые измерения Чтобы изучить RWC в ранней и поздней древесине, восемь доминирующих деревьев были взяты буром с шагом 12 мм зимой (10–15 марта 2001 г., перед вегетационным периодом) и летом (25–30 сентября 2001 г., перед началом вегетационного периода). начался дождь).Сердечники сразу после извлечения заворачивали в полиэтиленовую пленку и помещали в холодильник. В тот же день они были возвращены в лабораторию и обработаны. Последнее кольцо роста было удалено, а следующие два кольца роста были разделены на раннюю древесину и позднюю древесину. Как описано ранее, были измерены V f , M f и M d и рассчитано RWC . В те же дни и на тех же деревьях оценивали Ψ ствола на высоте груди (на листьях, завернутых в алюминиевую фольгу для предотвращения испарения) с помощью барокамеры (PMS Instrument Company, Корваллис, Орегон).Было измерено три лиственные ветви на дереве. Соотношение между отрицательным значением приложенного давления и RWC и PLC определяли по образцам, испытанным в лаборатории, затем оценивали PLC на основе измеренных Ψ и RWC стоящих деревьев в поле.
Последнее кольцо роста было удалено, а следующие два кольца роста были разделены на раннюю древесину и позднюю древесину. Как описано ранее, были измерены V f , M f и M d и рассчитано RWC . В те же дни и на тех же деревьях оценивали Ψ ствола на высоте груди (на листьях, завернутых в алюминиевую фольгу для предотвращения испарения) с помощью барокамеры (PMS Instrument Company, Корваллис, Орегон).Было измерено три лиственные ветви на дереве. Соотношение между отрицательным значением приложенного давления и RWC и PLC определяли по образцам, испытанным в лаборатории, затем оценивали PLC на основе измеренных Ψ и RWC стоящих деревьев в поле.
Статистический анализ
Метод наименьших квадратов использовался для подбора взаимосвязей между гидравлическими параметрами и приложенным давлением, а также линейных и нелинейных взаимосвязей между гидравлическими параметрами. Дисперсионный анализ (ANOVA) использовался для определения различий в гидравлических параметрах между целыми образцами и подмножеством образцов с деревом в качестве блокирующего фактора. Парные t ‐испытания использовались специально для проверки разницы в гидравлических параметрах между ранней и поздней древесиной. Все статистические процедуры проводились с использованием программного обеспечения Statistical Analysis Systems ((SAS Institute Inc, 1997).
Дисперсионный анализ (ANOVA) использовался для определения различий в гидравлических параметрах между целыми образцами и подмножеством образцов с деревом в качестве блокирующего фактора. Парные t ‐испытания использовались специально для проверки разницы в гидравлических параметрах между ранней и поздней древесиной. Все статистические процедуры проводились с использованием программного обеспечения Statistical Analysis Systems ((SAS Institute Inc, 1997).
Результаты
Форма образца, тесты окрашивания
Средняя поздняя древесина составила 33% кольца (±2%; колебалась от 26% до 40%) (табл. 1).Сумма измеренных k s в ранней древесине и поздней древесине, взвешенная по доле ранней/поздней древесины, была хорошей оценкой k s , измеренных в пределах целых образцов (рис. 3а). Наклон линии регрессии достоверно не отличался от 1 ( P =0,26). Ни гидравлические параметры, ни расчетные емкости не отличались между целыми образцами (Б-Ф) и частями подвыборки (В 1 , В 2 до Ф 1 , Ф 2 ; Р >0).62, данные не показаны). Целые длинные образцы (A) и небольшие части образца (D–F) имели аналогичные PLC при 0,5, 1,0 и 3,0 МПа (рис. 3б). Наблюдалась тесная взаимосвязь между оценкой PLC на основе неокрашенной части образцов (нефункциональная древесина) и фактическим измеренным PLC (рис. 4). Однако наклон линии регрессии был значительно ниже 1 ( P =0,04).
Ни гидравлические параметры, ни расчетные емкости не отличались между целыми образцами (Б-Ф) и частями подвыборки (В 1 , В 2 до Ф 1 , Ф 2 ; Р >0).62, данные не показаны). Целые длинные образцы (A) и небольшие части образца (D–F) имели аналогичные PLC при 0,5, 1,0 и 3,0 МПа (рис. 3б). Наблюдалась тесная взаимосвязь между оценкой PLC на основе неокрашенной части образцов (нефункциональная древесина) и фактическим измеренным PLC (рис. 4). Однако наклон линии регрессии был значительно ниже 1 ( P =0,04).
Гидравлические параметры ранней и поздней древесины
Ранняя древесина имела значительно более высокие значения k s , чем поздняя древесина ( P <0.001, табл. 2). Поздняя древесина содержала менее 9% k s ранней древесины и 13% k s цельной древесины. В кольцевой шкале поздняя древесина (составляющая 33 % площади поверхности) дала только 5 % от общего количества k h (табл. 2).
2).
Уязвимость к эмболии варьировала среди образцов древесины. Образцы поздней древесины были более уязвимы к эмболии, чем образцы ранней древесины при 0,5 МПа и 1,0 МПа, но не при 3,0 МПа (табл. 2; рис. 5а). Образцы ранней древесины существенно не отличались от образца цельной древесины ( P >0.21) и следовали той же тенденции. Давление ксилемы, вызывающее 50 PLC ( P 50 ) для всего VC и ранней древесины, составляло –2,3±0,1 и –2,1±0,2 МПа соответственно (рис. 5а). При любом приложенном давлении образцы цельной древесины и образцы ранней древесины значительно отличались от образцов поздней древесины ( P <0,04).
Водный дефицит (100– RWC ) показал сигмоидальное изменение с приложенным давлением в образцах цельной древесины. Самый крутой склон был между 2.0 и 3,0 МПа (рис. 5б). Для поздней древесины водный дефицит увеличивался экспоненциально с увеличением приложенного давления, достигая максимума при 40% (рис. 5b). Ранняя древесина имела меньшее снижение водного дефицита при 0,5 и 1,0 МПа, чем поздняя древесина, но гораздо большее снижение и значительно отличалась от поздней древесины при 3,0 МПа ( P =0,03). Образцы ранней древесины имели такой же дефицит воды, как и образцы цельной древесины при 0,5 и 1,0 МПа, но они имели значительно более низкие значения при 3,0 МПа ( P < 0.01). Поздняя древесина имела значительно отличающиеся значения водного дефицита, чем образцы цельной древесины для трех различных приложенных давлений ( P <0,05, рис. 5b).
5b). Ранняя древесина имела меньшее снижение водного дефицита при 0,5 и 1,0 МПа, чем поздняя древесина, но гораздо большее снижение и значительно отличалась от поздней древесины при 3,0 МПа ( P =0,03). Образцы ранней древесины имели такой же дефицит воды, как и образцы цельной древесины при 0,5 и 1,0 МПа, но они имели значительно более низкие значения при 3,0 МПа ( P < 0.01). Поздняя древесина имела значительно отличающиеся значения водного дефицита, чем образцы цельной древесины для трех различных приложенных давлений ( P <0,05, рис. 5b).
Поздняя древесина обладала гораздо более высокой водоаккумулирующей способностью, чем ранняя древесина, от 0,5 до 1,0 МПа (22 против 4% RWC МПа –1 , поздняя и ранняя древесина, соответственно). Эта тенденция менялась на противоположную в диапазоне от 1,0 до 3,0 МПа (5% по сравнению с 23% RWC МПа –1 для поздней и ранней древесины соответственно; таблица 3).Для C 0–0,5 существенной разницы между ранней и поздней древесиной не было (табл. 3). Для C 0,5–1,0 не было существенной разницы между цельным образцом и поздней древесиной, но была значительная разница между целым образцом и ранней древесиной и между целым образцом и поздней древесиной C 0–0,5 и C 1,0–3,0 ( P <0,05). В образцах ранней древесины наблюдалась значительная разница в емкости хранения воды между C 0–0.5, С 0,5–1,0 и С 1,0–3,0 ( Р <0,006). Влагоаккумулирующая способность между первыми двумя фазами для образцов поздней древесины ( P =0,9) и между двумя последними фазами для образцов цельной древесины ( P =0,7) существенно не различалась между первыми двумя фазами для образцов цельной древесины ( P =0,7)
3). Для C 0,5–1,0 не было существенной разницы между цельным образцом и поздней древесиной, но была значительная разница между целым образцом и ранней древесиной и между целым образцом и поздней древесиной C 0–0,5 и C 1,0–3,0 ( P <0,05). В образцах ранней древесины наблюдалась значительная разница в емкости хранения воды между C 0–0.5, С 0,5–1,0 и С 1,0–3,0 ( Р <0,006). Влагоаккумулирующая способность между первыми двумя фазами для образцов поздней древесины ( P =0,9) и между двумя последними фазами для образцов цельной древесины ( P =0,7) существенно не различалась между первыми двумя фазами для образцов цельной древесины ( P =0,7)
Состав поздней и ранней древесины
Процентное содержание воздуха и RWC при полном насыщении были одинаковыми для обоих типов древесины, однако все остальные свойства древесины статистически различались ( P <0. 01; Таблица 4). Поздняя древесина имела более высокий процент клеточной стенки и более низкий процент воды. При 3,0 МПа поздняя древесина имела более низкий процент воздуха, но такой же процент воды. Поздняя древесина имела более чем в два раза большую плотность, чем ранняя древесина (0,26 г см -3 против 0,60 г см -3 ).
01; Таблица 4). Поздняя древесина имела более высокий процент клеточной стенки и более низкий процент воды. При 3,0 МПа поздняя древесина имела более низкий процент воздуха, но такой же процент воды. Поздняя древесина имела более чем в два раза большую плотность, чем ранняя древесина (0,26 г см -3 против 0,60 г см -3 ).
Сравнение полевых измерений и лабораторных оценок
Минимальный ствол Ψ, измеренный в конце лета, никогда не опускался ниже –1,4±0,2 МПа, при расчетном минимуме RWC 69±3% для всего кольца.Поздняя древесина RWC была значительно ниже, чем ранняя древесина RWC ( P =0,002). Сезонное снижение RWC в ранней и поздней древесине составило 4% и 8% соответственно (таблица 5). Рассчитанные значения PLC из летнего ствола Ψ составили 16% и 42% в ранней и поздней древесине соответственно (табл. 5). Наблюдалось близкое соответствие между оценкой PLC с по измеренному Ψ и RWC для ранней древесины, но не для поздней древесины.
Обсуждение
Методологические вопросы
Использование небольших или длинных образцов дало статистически неразделимые результаты для k s или уязвимости к эмболии (рис.3б). Разделение образцов на две части (одну для ранней древесины, другую для поздней) не повлияло на степень эмболии, что позволило продолжить анализ данных. Это открытие показывает, что повреждение краев образца не оказывает существенного влияния на k s или уязвимость к эмболии, что подтверждает пригодность этого метода (Domec and Gartner, 2001).
Сегментация ранней/поздней древесины по проводимости и подверженности эмболии
Как и ожидалось, k s у поздней древесины намного ниже, чем у ранней древесины.Очень низкий k s , обнаруженный в поздней древесине, можно объяснить малым диаметром трахеид ее клеток по сравнению с клетками ранней древесины. Закон Хагена-Пуазейля предсказывает, что большие трахеиды ранней древесины отвечают за большую часть потока воды в пределах одного кольца (Zimmermann, 1983). Ранняя древесина k s превышала позднюю древесину k s в 11 раз, что соответствовало бы увеличению диаметра трахеи в 1,8 раза (закон Гагена-Пуазейля) при прочих равных условиях.Этот результат согласуется с описанным в литературе соотношением диаметров трахеид для хвойных (МсМи1ап, 1968; Соуп, 1975).
Закон Хагена-Пуазейля предсказывает, что большие трахеиды ранней древесины отвечают за большую часть потока воды в пределах одного кольца (Zimmermann, 1983). Ранняя древесина k s превышала позднюю древесину k s в 11 раз, что соответствовало бы увеличению диаметра трахеи в 1,8 раза (закон Гагена-Пуазейля) при прочих равных условиях.Этот результат согласуется с описанным в литературе соотношением диаметров трахеид для хвойных (МсМи1ап, 1968; Соуп, 1975).
Результаты подтвердили гипотезу о том, что ранняя древесина более эффективно транспортирует воду и менее подвержена эмболии, чем поздняя древесина при Ψ, встречающемся в полевых условиях в нормальных условиях (табл. 2, 5). Этот результат контрастирует с полученным для кольцевидных видов ((Lo Gullo et al. ., 1995). Используя метод окрашивания, они показали, что сосуды поздней древесины молодых деревьев Quercus cerris были более узкими, короткими и менее уязвимыми. к эмболии, чем ранние.
к эмболии, чем ранние.
Внутри кольца роста наблюдалась положительная связь между k s и безопасностью, и между ними не было компромисса. Однако запас прочности между измеренным Ψ в поле и началом эмболии был менее 0,5 МПа. Это исследование подтвердило, что древесина основания ствола пихты Дугласа может работать на грани гидравлической безопасности из-за высокой уязвимости ранней древесины к эмболии ниже –2,0 МПа ((Domec and Gartner, 2002 a ).
Считается, что образование эмболии связано не с размером кондуита, а скорее с размером пор на границе раздела двух трахеид через окаймленные ямки (Tyree et al. ., 1994). Действительно, низкая устойчивость к эмболии коррелирует со структурными особенностями мембраны окаймленной ямки (Petty, 1972; Bolton and Petty, 1978; (Domec, Gartner, 2002 b ). обнаруженный в поздней древесине, может не позволить марго действовать как клапан, предотвращая распространение пузырьков воздуха от одной эмболизированной трахеиды к другой. Сообщалось, что у Pinus sylvestris (L.) мембраны поздней древесины были более жесткими, чем мембраны ранней древесины, и что они не могли полностью изолироваться при значениях поля Ψ (Gregory, Petty, 1973). Поры между нитями мембраны в поздней древесине меньше, чем в ранней древесине (Petty and Puritch, 1970; Fengel, 1972), что позволяет предположить, что засев воздуха происходит непосредственно через эти поры. Механическое сопротивление тора превысит механическое напряжение, необходимое для протягивания пузырька воздуха через мембрану.Тогда механизм эмболии, вызванной водным стрессом, у поздней древесины будет подобен механизму у несущих сосуды растений (Sperry and Tyree, 1988). Тогда устойчивость поздней древесины к эмболии будет пропорциональна размеру пор в поздней древесине, что может объяснить наблюдаемое снижение проводимости, начинающееся сразу при 0 МПа и ниже. Форма кривой до выхода на плато (0–1,0 МПа, рис. 5б) может отражать распределение пор по размерам в нитях мембраны поздней древесины.
Сообщалось, что у Pinus sylvestris (L.) мембраны поздней древесины были более жесткими, чем мембраны ранней древесины, и что они не могли полностью изолироваться при значениях поля Ψ (Gregory, Petty, 1973). Поры между нитями мембраны в поздней древесине меньше, чем в ранней древесине (Petty and Puritch, 1970; Fengel, 1972), что позволяет предположить, что засев воздуха происходит непосредственно через эти поры. Механическое сопротивление тора превысит механическое напряжение, необходимое для протягивания пузырька воздуха через мембрану.Тогда механизм эмболии, вызванной водным стрессом, у поздней древесины будет подобен механизму у несущих сосуды растений (Sperry and Tyree, 1988). Тогда устойчивость поздней древесины к эмболии будет пропорциональна размеру пор в поздней древесине, что может объяснить наблюдаемое снижение проводимости, начинающееся сразу при 0 МПа и ниже. Форма кривой до выхода на плато (0–1,0 МПа, рис. 5б) может отражать распределение пор по размерам в нитях мембраны поздней древесины. Мембраны, вероятно, полностью прогибались при Ψ <–1,0 МПа, и герметичный тор, вероятно, создавал замкнутую систему, защищающую трахеиды, ответственные за оставшиеся 40% проводимости. Относительно ранней древесины сообщалось, что пористость мембраны, удерживающей тор, слишком велика, чтобы предотвратить прохождение воздушного мениска при значениях Ψ ниже –0,1 МПа, но она достаточно эластична, чтобы полностью прогибаться и герметизировать тор (Сперри и Тайри, 1990). Соскальзывание тора из его закрытого положения создало зазоры, достаточно большие для возникновения кавитации в соседних трахеидах.Этот результат может объяснить, почему была полная потеря проводимости при более низких приложенных давлениях в ранней древесине, но не в поздней древесине (рис. 5а). Более толстый тор поздней древесины предотвратил бы его соскальзывание с закрытого положения (Domec and Gartner, 2002 b ).
Мембраны, вероятно, полностью прогибались при Ψ <–1,0 МПа, и герметичный тор, вероятно, создавал замкнутую систему, защищающую трахеиды, ответственные за оставшиеся 40% проводимости. Относительно ранней древесины сообщалось, что пористость мембраны, удерживающей тор, слишком велика, чтобы предотвратить прохождение воздушного мениска при значениях Ψ ниже –0,1 МПа, но она достаточно эластична, чтобы полностью прогибаться и герметизировать тор (Сперри и Тайри, 1990). Соскальзывание тора из его закрытого положения создало зазоры, достаточно большие для возникновения кавитации в соседних трахеидах.Этот результат может объяснить, почему была полная потеря проводимости при более низких приложенных давлениях в ранней древесине, но не в поздней древесине (рис. 5а). Более толстый тор поздней древесины предотвратил бы его соскальзывание с закрытого положения (Domec and Gartner, 2002 b ).
Обратный расчет доли поздней древесины в остальных k s при –3,0 МПа (рис.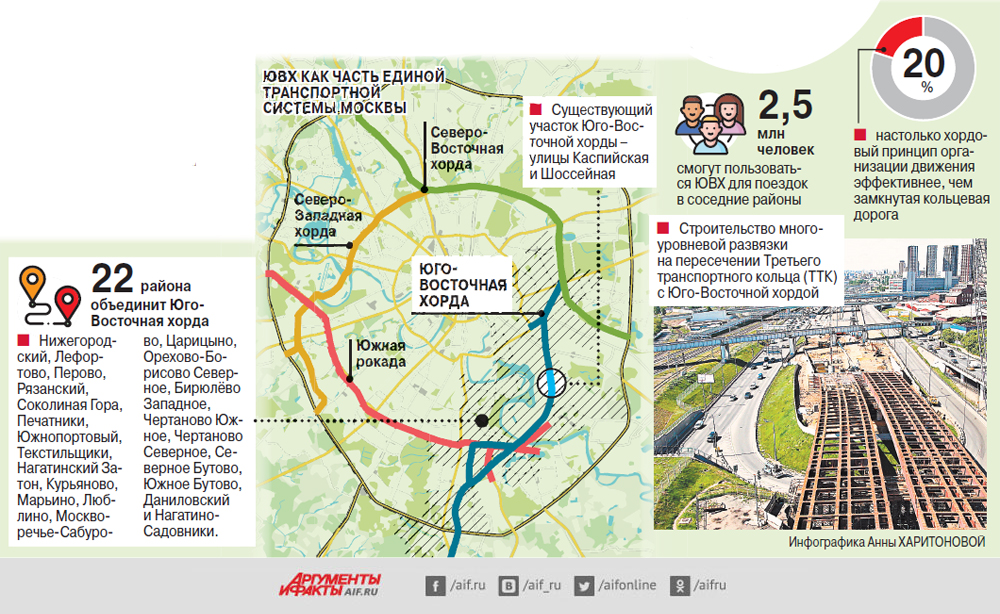 5а) показал, что незапечатывание ямок, окаймленных поздней древесиной, у пихты Дугласа, по-видимому, составляет 30% от общего оставшаяся гидропроводность.Однако при очень низком k s и доле (33%) в кольце полностью проводящая поздняя древесина может составлять только 5% начальной проводимости при полном насыщении, хотя на нее может приходиться 16% оставшейся проводимости. при –3,0 МПа. Эта остаточная электропроводность поздней древесины составляет всего 2% от исходной электропроводности, что далеко от 20%, рассчитанных Сперри и Тайри (1990). Взяв взрослое дерево с небольшими годичными кольцами и более высокой долей поздней древесины (около 55%) (De Kort, 1993), поздняя древесина по-прежнему будет составлять только 11% от общей проводимости кольца при –3.0 МПа (при сильной засухе). Чтобы обеспечить 50% проводимости всего кольца, поздняя древесина должна составлять 90% всей площади кольца при полном насыщении и 75% при -3,0 МПа. Такая высокая доля поздней древесины в годичном кольце вряд ли может быть обнаружена в древесине обычных хвойных пород, но она часто встречается в сжатой древесине, которая образуется на нижних сторонах ветвей или в наклонных стволах (Westing, 1968; Timell, 1986).
5а) показал, что незапечатывание ямок, окаймленных поздней древесиной, у пихты Дугласа, по-видимому, составляет 30% от общего оставшаяся гидропроводность.Однако при очень низком k s и доле (33%) в кольце полностью проводящая поздняя древесина может составлять только 5% начальной проводимости при полном насыщении, хотя на нее может приходиться 16% оставшейся проводимости. при –3,0 МПа. Эта остаточная электропроводность поздней древесины составляет всего 2% от исходной электропроводности, что далеко от 20%, рассчитанных Сперри и Тайри (1990). Взяв взрослое дерево с небольшими годичными кольцами и более высокой долей поздней древесины (около 55%) (De Kort, 1993), поздняя древесина по-прежнему будет составлять только 11% от общей проводимости кольца при –3.0 МПа (при сильной засухе). Чтобы обеспечить 50% проводимости всего кольца, поздняя древесина должна составлять 90% всей площади кольца при полном насыщении и 75% при -3,0 МПа. Такая высокая доля поздней древесины в годичном кольце вряд ли может быть обнаружена в древесине обычных хвойных пород, но она часто встречается в сжатой древесине, которая образуется на нижних сторонах ветвей или в наклонных стволах (Westing, 1968; Timell, 1986). .
.
Сжатая древесина стволов имеет широкие годовые кольца и намного плотнее обычной древесины из-за повышенной доли поздней древесины (Park, 1984; Yoshizawa et al ., 1987). Было показано, что сжатая древесина ветвей и сеянцев пихты Дугласа имеет значительно более низкие значения k s , чем нормальная древесина (Spicer and Gartner, 1998 b ). Можно предположить, что компрессионная древесина в стволах будет более уязвима для эмболии, чем обычная древесина при высоких значениях Ψ. С другой стороны, она может быть более устойчивой к сильной засухе из-за более высокой устойчивости поздней древесины к эмболии при низком Ψ. Старые деревья также обычно имеют более высокую долю поздней древесины, чем молодые или взрослые деревья, что может ограничивать их перенос воды, но повышает их устойчивость к эмболии при очень низком Ψ.Более высокое соотношение поздней и ранней древесины может быть стратегией для таких видов, как пихта Дугласа, растущих во влажных условиях весной и в сухих условиях летом. Это также может объяснить, почему уязвимость к эмболии у взрослых деревьев пихты Дугласа была выше у основания ствола, чем в верхней части ствола (под кроной), тогда как ее k s была ниже (Domec and Gartner, 2001).
Это также может объяснить, почему уязвимость к эмболии у взрослых деревьев пихты Дугласа была выше у основания ствола, чем в верхней части ствола (под кроной), тогда как ее k s была ниже (Domec and Gartner, 2001).
Различия в емкости хранения воды между ранней и поздней древесиной
Высокая водоаккумулирующая способность, наблюдаемая при высоком Ψ (–0.5 МПа) описывается как капиллярная вода (Tyree and Yang, 1990) и не имеет адаптивного значения, поскольку возникает до того, как растение испытывает водный стресс. Поэтому, хотя первые значения емкости не были объяснены между 0 и 0,5 МПа, два других расчетных значения емкости (Ψ = 0,5–1,0 и 1,0–3,0 МПа) оказались подходящими.
Исследование, описанное в этой статье, показало, что в годичном кольце поздняя древесина обладает более высокой влагоемкостью, чем ранняя древесина (рис. 5b), и более уязвима для эмболии в поле Ψ в нормальных условиях (>–2.0 МПа). Для ранней древесины низкая аккумулирующая способность, обнаруженная для рабочего диапазона потенциалов (между 0–1 МПа), предполагает, что ее водопоглощение будет линейно связано с потерей воды листьями (табл. 3; рис. 5б). Преимущество поддержания этого стационарного потока подчеркивается тем фактом, что мало случаев эмболии происходит при Ψ менее отрицательном, чем -2,0 МПа (рис. 5а). Таким образом, для ранней древесины компромиссом, связанным с низкой уязвимостью к эмболии и высокой k s , является низкая влагоемкость (Goldstein et al ., 1998; Stratton и др. ., 2000). Эти характеристики могут быть важны для старых деревьев с более высоким процентом поздней древесины, и можно предположить, что для естественного диапазона Ψ ранняя древесина лучше всего будет служить дереву, эффективно и безопасно проводя воду, а поздняя древесина, обеспечивая значительное количество запасенной воды. . С другой стороны, в сухих условиях (< –2,0 МПа) ранняя древесина с большей вероятностью, чем поздняя древесина, будет использовать накопление воды, которое может представлять собой важную фракцию, но тогда поздняя древесина будет поддерживать минимальный уровень k s за счет лучшего устойчивость к эмболии.
3; рис. 5б). Преимущество поддержания этого стационарного потока подчеркивается тем фактом, что мало случаев эмболии происходит при Ψ менее отрицательном, чем -2,0 МПа (рис. 5а). Таким образом, для ранней древесины компромиссом, связанным с низкой уязвимостью к эмболии и высокой k s , является низкая влагоемкость (Goldstein et al ., 1998; Stratton и др. ., 2000). Эти характеристики могут быть важны для старых деревьев с более высоким процентом поздней древесины, и можно предположить, что для естественного диапазона Ψ ранняя древесина лучше всего будет служить дереву, эффективно и безопасно проводя воду, а поздняя древесина, обеспечивая значительное количество запасенной воды. . С другой стороны, в сухих условиях (< –2,0 МПа) ранняя древесина с большей вероятностью, чем поздняя древесина, будет использовать накопление воды, которое может представлять собой важную фракцию, но тогда поздняя древесина будет поддерживать минимальный уровень k s за счет лучшего устойчивость к эмболии. Эта гипотеза могла бы объяснить, почему разница между RWC в поздней древесине была больше, чем в ранней древесине между зимой и летом (таблица 5). Эта гипотеза также может объяснить, почему деревья с более высокой плотностью древесины и более высоким процентом поздней древесины относятся к засушливым районам (Polge, 1973; Megraw, 1985; Barajas-Morales, 1985) и более устойчивы к кавитации (Alder et al ., 1996). ; Hacke и др. , 2001).
Эта гипотеза могла бы объяснить, почему разница между RWC в поздней древесине была больше, чем в ранней древесине между зимой и летом (таблица 5). Эта гипотеза также может объяснить, почему деревья с более высокой плотностью древесины и более высоким процентом поздней древесины относятся к засушливым районам (Polge, 1973; Megraw, 1985; Barajas-Morales, 1985) и более устойчивы к кавитации (Alder et al ., 1996). ; Hacke и др. , 2001).
Компромисс между гидравлическими и механическими свойствами поздней и ранней древесины
Поздний лес имел плотность 2.в 3 раза выше, чем в ранней древесине (табл. 4), аналогично другим данным для того же вида (Lutz, 1964; (Takjima, 1967). Завышенным фактором, определяющим прочность древесины среди деревьев, является различное процентное содержание поздней древесины для большинства видов (Zobel and Van). Buijtenen, 1989). Деревья пихты Дугласа растут в основном в весеннее время, когда много дождя и солнца, поддерживающих рост (Waring and Running, 1998)..jpg) Переход от ранней древесины к поздней тесно связан с наличием влаги в почве. , потому что стресс от высокой влажности снижал выработку ауксина (Gilmore et al ., 1966). Регуляция диаметра клеток является гормональной и опосредуется ауксином, образующимся в почках растущих побегов. Высокое производство ауксина запускает производство ранней древесины (рассмотрено в Larson, 1994; Aloni, 2001).
Переход от ранней древесины к поздней тесно связан с наличием влаги в почве. , потому что стресс от высокой влажности снижал выработку ауксина (Gilmore et al ., 1966). Регуляция диаметра клеток является гормональной и опосредуется ауксином, образующимся в почках растущих побегов. Высокое производство ауксина запускает производство ранней древесины (рассмотрено в Larson, 1994; Aloni, 2001).
Для данного количества углерода, хранящегося в виде заболони, изменение соотношения поздней и ранней древесины в годичных кольцах с 0,5 (соотношение, обнаруженное у исследованных деревьев) до 2 всего за 1 год увеличило бы общую плотность древесины на 34%, что также дают увеличение модуля Юнга ( E или жесткость древесины) в том же диапазоне (Biblis, 1969; Mott et al )., 2002), поскольку плотность древесины и E коррелируют с коэффициентом, близким к 1 (Bodig and Jayne, 1982). Механические характеристики (судя по структурной жесткости) значительно возрастают по мере увеличения диаметра образца, но это изменение определяется увеличением второго момента площади ( I , масштаб которого зависит от радиуса стержня в четвертой степени). что почти полностью стирает вклад E . Ключевое частное будет тогда E × I (структурная жесткость), и изменение отношения поздней/ранней древесины до 2 увеличит E × I только на 26% (при средней скорости роста 8.6 мм год –1 , таблица 1), потому что для данного количества хранимой древесины более высокое соотношение поздней и ранней древесины также будет означать более узкое годичное кольцо. С другой стороны, k s уменьшится на 46 %, но общая заболонь C 0,5–1,0 увеличится на 55 %. Таким образом, изменение соотношения поздняя/ранняя древесина всего за 1 год окажет большее влияние на гидравлические параметры, чем на механическую стойкость заболони. Компромисс и преимущество заключались бы в том, что из-за увеличения емкости деревьям было бы доступно больше накопленной воды.
что почти полностью стирает вклад E . Ключевое частное будет тогда E × I (структурная жесткость), и изменение отношения поздней/ранней древесины до 2 увеличит E × I только на 26% (при средней скорости роста 8.6 мм год –1 , таблица 1), потому что для данного количества хранимой древесины более высокое соотношение поздней и ранней древесины также будет означать более узкое годичное кольцо. С другой стороны, k s уменьшится на 46 %, но общая заболонь C 0,5–1,0 увеличится на 55 %. Таким образом, изменение соотношения поздняя/ранняя древесина всего за 1 год окажет большее влияние на гидравлические параметры, чем на механическую стойкость заболони. Компромисс и преимущество заключались бы в том, что из-за увеличения емкости деревьям было бы доступно больше накопленной воды.
Благодарности
Этот проект был поддержан USDA CSREES 97-35103-5052, а также специальным грантом USDA Университету штата Орегон на исследования в области использования древесины. Авторы благодарят Ричарда Саймонса, Эндрю Чанга и Дункана Уилсона за помощь в вырубке деревьев в Макдональдс Форест. Мы также благодарим Марселя Мерка, Дженнифер Свенсон, Лорана Буффье и Рэнди Джонсона за помощь на местах, а также Б. Дж. Бонда, С. Дж. Кристи, М. Ансуорта, Г. Хоу и Р. Лейхти за исправления рукописи.Это документ 3533 Лаборатории лесных исследований Университета штата Орегон, Корваллис, штат Орегон.
Авторы благодарят Ричарда Саймонса, Эндрю Чанга и Дункана Уилсона за помощь в вырубке деревьев в Макдональдс Форест. Мы также благодарим Марселя Мерка, Дженнифер Свенсон, Лорана Буффье и Рэнди Джонсона за помощь на местах, а также Б. Дж. Бонда, С. Дж. Кристи, М. Ансуорта, Г. Хоу и Р. Лейхти за исправления рукописи.Это документ 3533 Лаборатории лесных исследований Университета штата Орегон, Корваллис, штат Орегон.
Рис. 1. Пробоподготовка. Шесть образцов в пределах колец 3–7 были взяты из диска дерева, взятого на высоте груди. Из шести подготовленных образцов один образец (образец А) случайным образом использовали для определения гидравлических свойств всего кольца («цельная древесина»). Остальные пять образцов (образцы B–F) были разделены на два равных сегмента для определения гидравлических свойств ранней и поздней древесины (B 1 , B 2 … F 1 , F 2 ).Для ясности показан только образец B.
Рис. 1. Пробоподготовка. Шесть образцов в пределах колец 3–7 были взяты из диска дерева, взятого на высоте груди. Из шести подготовленных образцов один образец (образец А) случайным образом использовали для определения гидравлических свойств всего кольца («цельная древесина»). Остальные пять образцов (образцы B–F) были разделены на два равных сегмента для определения гидравлических свойств ранней и поздней древесины (B 1 , B 2 … F 1 , F 2 ).Для ясности показан только образец B.
1. Пробоподготовка. Шесть образцов в пределах колец 3–7 были взяты из диска дерева, взятого на высоте груди. Из шести подготовленных образцов один образец (образец А) случайным образом использовали для определения гидравлических свойств всего кольца («цельная древесина»). Остальные пять образцов (образцы B–F) были разделены на два равных сегмента для определения гидравлических свойств ранней и поздней древесины (B 1 , B 2 … F 1 , F 2 ).Для ясности показан только образец B.
Рис. 2. Краткое описание процедур, использованных для анализа удельной электропроводности ( k s ) и кривых уязвимости в пределах одного кольца или всего образца (A), а также образцов ранней и поздней древесины (B–F).
Рис. 2. Краткое описание процедур, использованных для определения удельной электропроводности ( k s ) и кривых уязвимости в пределах одного кольца или всего образца (A), а также образцов ранней и поздней древесины (B–F).
Рис. 3. (а) Измеренная удельная электропроводность ( к с ) во всем образце по сравнению с к с оценена как сумма измеренных к с 90 взвешенной древесины соотношением ранней и поздней древесины в кольце ( n = 20). Представлены линия регрессии и 95% доверительные интервалы. Наклон линии регрессии достоверно не отличался от 1 ( P =0.26). (b) Сравнение процентной потери проводимости ( PLC ) при трех приложенных давлениях между маленькими и целыми образцами.
Рис. 3. (a) Измеренная удельная электропроводность ( k с ) во всем образце по сравнению с k s рассчитана как сумма измеренных k с в ранней древесине и поздней древесине взвешено по соотношению ранней и поздней древесины в кольце ( n = 20). Представлены линия регрессии и 95% доверительные интервалы.Наклон линии регрессии достоверно не отличался от 1 ( P =0,26). (b) Сравнение процентной потери проводимости ( PLC ) при трех приложенных давлениях между маленькими и целыми образцами.
(b) Сравнение процентной потери проводимости ( PLC ) при трех приложенных давлениях между маленькими и целыми образцами.
Рис. 4. Эмболизированные трахеиды, измеренные по проценту потери проводимости ( PLC , уравнение 3) во всем образце и в подобразцах ранней и поздней древесины в сравнении с площадью эмболизации, оцененной методом окрашивания (% неокрашенной площади).Представлены линия регрессии и 95% доверительные интервалы ( n =80). Наклон линии регрессии значительно отличался от 1 ( P =0,04).
Рис. 4. Эмболизированные трахеиды, измеренные по потере проводимости в процентах ( PLC , уравнение 3) во всем образце и в подобразцах ранней и поздней древесины в сравнении с площадью эмболизации, оцененной методом окрашивания (% неокрашенной площади). Представлены линия регрессии и 95% доверительные интервалы ( n =80).Наклон линии регрессии значительно отличался от 1 ( P =0,04).
Рис. 5. (a) Кривые уязвимости, показывающие процент потери гидравлической проводимости ксилемы ( PLC ) и (b) дефицит воды (100– RWC ) для цельной, ранней и поздней древесины пихты Дугласа 21-летних деревьев на высоте груди по сравнению с отрицательным давлением воздуха, используемым в экспериментах по нагнетанию воздуха. Столбики погрешностей представляют собой стандартные ошибки ( n =5).
Рис. 5. (a) Кривые уязвимости, показывающие процентную потерю гидравлической проводимости ксилемы ( PLC ) и (b) дефицит воды (100– RWC ) для цельной древесины пихты Дугласа, ранней древесины и поздняя древесина 21-летних деревьев на высоте груди по сравнению с отрицательным давлением воздуха, используемым в экспериментах с нагнетанием воздуха. Столбики погрешностей представляют собой стандартные ошибки ( n =5).
Таблица 1.Дерево и образцы Характеристики Изученных Дугласа-елей ( N = 5)
| Среднее ± SE | Minime | максимум | ||
| Возраст дерева (лет) | 21. 4 ± 0,5 4 ± 0,5 | 23 | 23 | 20 |
| Высота дерева (M) | 17.4 ± 0,6 | 20 | 16 | |
| Дерево-недвижимость на высоте (высота груди) | ||||
| Возраст дерева (лет) | 17.5 ± 0,6 | 15.2 | 18.8 | |
| 29,4 ± 0,8 | 28 | 31.7 | ||
| Sapwood (см 2 ) | 424 ± 31 | 374 518 | ||
| Heartwood области (см 2 | ) 176 ± 10 | 133 | 199 | |
| Количество заболони кольца | 9.4 ± 0,5 | 8 | 11 | |
| свойства Кольцо | ||||
| кольцо из коры использовали | 4,4 ± 0,6 | 3 | 7 | |
| Кольцо (мм) | 8,6 ± 0,3 | 9,2 ± 0,3 | 7,4 ± 0,3 | |
| Процент earlywood | 66,6 ± 2,3 | 59,7 ± 1,9 | 74,5 ± 1,9 | |
| Среднее ± SE | минимум | максимум | |||
| Свойства дерева | |||||
| Tree Age (лет) | 21. 4 ± 0,5 4 ± 0,5 | 23 | 23 | 20 | |
| Высота дерева (M) | 17.4 ± 0,6 | 20 | 16 | ||
| Дерево-недвижимость на высоте (высота груди) | |||||
| Возраст дерева (лет) | 17.5 ± 0,6 | 15.2 | 18.8 | ||
| 29,4 ± 0,8 | 28 | 31.7 | |||
| Sapwood (см 2 ) | 424 ± 31 | 374 518 | |||
| Heartwood области (см 2 | ) 176 ± 10 | 133 | 199 | ||
| Количество заболони кольца | 9.4 ± 0,5 | 8 | 11 | ||
| свойства Кольцо | |||||
| кольцо из коры использовали | 4,4 ± 0,6 | 3 | 7 | ||
| Кольцо (mm) | 81085 | 8,6 ± 0,3 | 9. 2 ± 0,3 2 ± 0,3 | 7,4 ± 0,3 | |
| Процент раннего рандения | 66,6 ± 2.3 | 59,7 ± 1,9 | 74,5 ± 1,9 | ||
Дерево и образцы Характеристики Дугласа-Еров ( N = 5)
| Среднее ± SE | минимум | максимум | |||
| Возраст дерева (лет) | 21.4 ± 0.5 | 23 | 20 | ||
| Высота дерева (M) | 17,4 ± 0,6 | 20 | 16 | ||
| Дерево высота выборки (высота груди) | |||||
| Возраст дерева (лет) | 17.5 ± 0.6 | 15.2 | 15.2 | 18.8 | |
| 29,4 ± 0.8 | 28 | 31. 7 7 | |||
| Sapwood Perlow (CM 2 ) | 424 ± 31 | 374 | 374 | 374 | 518 |
| Heartwood Sharewood (CM 2 ) | 176 ± 10 | 199 | |||
| Количество SASWOOD Rings | 9,4 ± 0.5 | 8 | 11 | ||
| Свойства кольца | |||||
| Кольцо из коры б/у | 4.4 ± 0,6 | 3 | 7 | 7 | |
| Кольцо ширина (мм) | 8,6 ± 0,3 | 9,2 ± 0,3 | 7,4 ± 0,3 | ||
| процент раннего дерева | 66,6 ± 2,3 | 59,7 ± 1,9 | 74.5 ± 1,9 | ||
| Minument ± SE | Минимум | максимум | ||
| Дерево свойства | ||||
| Tree Age (лет) | 21. 4 ± 0,5 4 ± 0,5 | 23 | 23 | 20 |
| Высота дерева (M) | 17.4 ± 0,6 | 20 | 16 | |
| Дерево-недвижимость на высоте (высота груди) | ||||
| Возраст дерева (лет) | 17.5 ± 0,6 | 15.2 | 18.8 | |
| 29,4 ± 0,8 | 28 | 31.7 | ||
| Sapwood (см 2 ) | 424 ± 31 | 374 518 | ||
| Heartwood области (см 2 | ) 176 ± 10 | 133 | 199 | |
| Количество заболони кольца | 9.4 ± 0,5 | 8 | 11 | |
| свойства Кольцо | ||||
| кольцо из коры использовали | 4,4 ± 0,6 | 3 | 7 | |
| Кольцо (мм) | 8.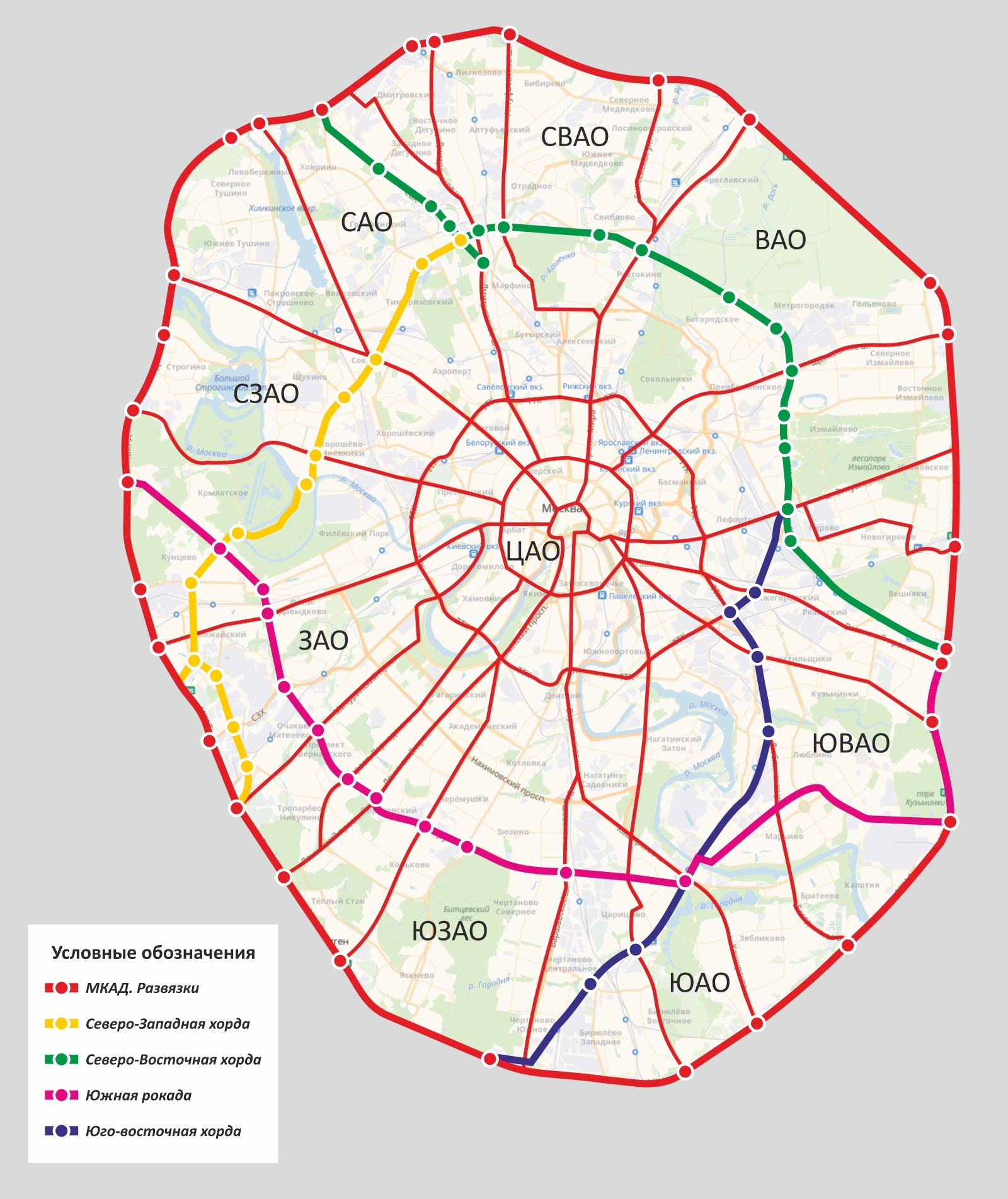 6 ± 0.3 6 ± 0.3 | 9.2 ± 0,3 | 7,4 ± 0,3 | |
| процент раннего дерева | 66,6 ± 2.3 | 59,7 ± 1,9 | 74,5 ± 1,9 | |
Удельная электропроводность ( k s ), гидравлическая проводимость ( k h ) и процентная потеря электропроводности ( PLC и образцы поздней древесины Среднее ±SE ( n =5). Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05.
| K сек (м 2 ) | к ч (м 4 ) | ПЛК (%) | ||||
| –0.5 МПа | -1.0 MPA | -3.0 MPA | ||||
| 5092 | ||||||
| 5,0 ± 0,3 × 10 -12 A | 3,8 ± 0,3 × 10 -16 A | 2..jpg) 9 ± 1,7 A 9 ± 1,7 A | 12.8 ± 4.5 A | 84.1 ± 1,1 A | ||
| Ранчание | 7,3 ± 0,5 × 10 -12 B | 3,4 ± 0,1 × 10 -16 A | 2,1 ± 1,9 A | 10.3 ± 6,4 | 90,2±2,8 | |
| Лейтвуд | 6.4 ± 0,9 × 10 -13 C | 2,0 ± 0,5 × 10 -17 A | 26,5 ± 9.1 b | 32.1 ± 8,3 b | 56,8 ± 89 | |
| K S (M 2 ) | K H (M 4 ) | PLC (%) | ||||
| -0,5 МПа | — 1,0 МПа | –3,0 МПа | ||||
| Цельная древесина | 5.0 ± 0,3 × 10 -12 A | 3,8 ± 0,3 × 10 -16 A | 2,9 ± 1,7 ± 4,5 A | 12,8 ± 4,5 A | 84.1 ± 1,1 A | |
| Ранчание | 7,3 ± 0,5 × 10 -12 B | 3. 4 ± 0,1 × 10 -16 A 4 ± 0,1 × 10 -16 A | 2,1 ± 1,9 A | 10,3 ± 6.4 A | 90,2 ± 2,8 A | |
| Railwood | 6,4 ± 0,9 × 10 –13 в | 2,0±0,5×10 –17 а | 26,5±9,1 б | 32.1 ± 8,3 b | 56,8 ± 8 | 56,8 ± 8.9 B |
( K S ), гидравлическая проводимость ( K H ) и процентная потеря электропроводность ( PLC ) при трех отрицательных значениях приложенного давления в образцах цельной древесины, ранней и поздней древесины Среднее ±SE ( n =5). Значения в столбце с одной и той же буквой существенно не различаются при P =0.05.
| K S (M S (M 2 ) | K H (M 4 ) | PLC (%) | ||||
-0.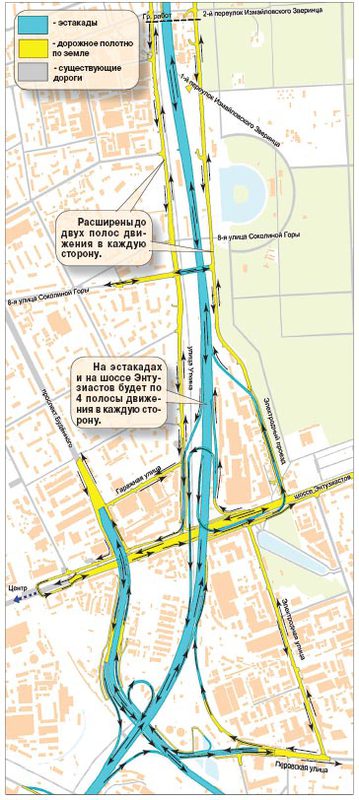 5 MPA 5 MPA | -1,0 МПа | -3.0 MPA | ||||
| 5,0 ± 0,3 × 10 | 5,0 ± 0,3 × 10 -12 A | 3,8 ± 0,3 × 10 -16 A | 2.9 ±1,7 г | 12.8 ± 4.5 A | 84.1 ± 1.1 A | |
| Ранчание | ||||||
| 7,3 ± 0,5 × 10 | 7,3 ± 0,5 × 10 -12 B | 3,4 ± 0,1 × 10 -16 A | 2,1 ± 1,9 A | 10,3 ± 6.4 | 90,2 ± 2.8 A | |
| 61084 | 6,4 ± 0,9 × 10 -13 C | 2,0 ± 0,5 × 10 | 2,0-17 A | 26,5 ± 9,1 млрд. | 32,1 ± 8,3 b | 56.8 ± 8,9 b |
| K S (M 2 (M 2 ) | K H (M 4 ) | PLC (%) | |||
| –0.5 МПа | -1.0 MPA | -3. 0 MPA 0 MPA | |||
| 5092 | |||||
| 5,0 ± 0,3 × 10 -12 A | 3,8 ± 0,3 × 10 -16 A | 2.9 ± 1,7 A | 12.8 ± 4.5 A | 84.1 ± 1,1 A | |
| Ранчание | 7,3 ± 0,5 × 10 -12 B | 3,4 ± 0,1 × 10 -16 A | 2,1 ± 1,9 A | 10.3 ± 6,4 | 90,2±2,8 |
| Лейтвуд | 6.4 ± 0,9 × 10 -13 C | 2,0 ± 0,5 × 10 -17 A | 26,5 ± 9.1 B | 32.1 ± 8,3 B | 56,8 ± 8,9 B |
Влияние типа древесины на емкость на основе RWC (емкость для хранения воды, % МПа –1 ) для трех фаз C 0–0,5
),
C 0,5–1,0 (от 0,5 до 1,0).0 МПа) и C 1,0–3,0 (от 1,0 до 3,0 МПа) Показаны средние значения (±SE) ( n =5). Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05.
Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05. C 0-0.5 | | 0.5-1.0 | | 5 C 1.0-3.0 | ||
| Цел-дерева | 5.1 ± 1,2 A | 11.3 ± 0,1 A | 15,3 ± 0,2 А | ||
| Ранчание | |||||
| 11,3 ± 4,3 A | 4,3 A | 4,1 ± 2,5 млн. | 23,5 ± 1,1 A | ||
| Латение | 10,5 ± 8,3 A | 21,8 ± 7.1 A | 5,5 ± 1,4 б |
| С 0-0,5 | С 0.5-1.0 | С 1,0-3,0 | |||
| общеправительственного дерево | 5.1 ± 1.2 A | 11,3 ± 0,1 A | 15,3 ± 0,2 | ||
| Ранчание | |||||
| 11,3 ± 4.3 A | 4,1 ± 2,5 млн. | 23,5 ± 1,1 A | |||
| Латение | 10,5 ± 8,3 а | 21,8 ± 7.1 A | 5,5 ± 1,4 млн. | 5,5 ± 1,4 млн. |
Эффект типа древесины на RWC -базируемых емкости (емкость хранения воды,% MPA -1 ) для трех фаз C 0–0,5 (от 0 до 0.5 МПа), C C 0.5-1.0 (от 0,5 до 1,0 МПа) и C 1.0-3.0 (от 1,0 до 3,0 МПа) означает (± SE) ( n =5). Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05.
| C 0-0.59 | 0.5-1.0 | C 1.0-3.0 | ||||
| Цел-лес | 5.1 ± 1.2 A | 11,3 ± 0,1 A | 15,3 ± 0,2 | |||
| Ранчание | ||||||
11,3 ± 4. 3 A 3 A | 4,1 ± 2,5 млн. | 23,5 ± 1,1 A | ||||
| Латение | 10,5 ± 8,3 а | 21,8 ± 7.1 | 5,5 ± 1,4 б |
| С 0-0,5 | С 0.5-1.0 | С 1,0-3,0 | |||
| Цельная древесина | 5.1 ± 1.2 A | 11,3 ± 0,1 A | 15,3 ± 0,2 | ||
| Ранчание | |||||
| 11,3 ± 4.3 A | 4,1 ± 2,5 млн. | 23,5 ± 1,1 A | |||
| Латение | 10,5 ± 8,3 а | 21,8±7,1 а | 5,5±1,4 б |
Плотность древесины (сухая масса / сырой объем), влажность ( MC ), объем, занимаемый материалом клеточной стенки ( V объем клеточной стенки ), занимаемый водой ( V H 2 ), Объем, занимаемый газом ( V ГАЗ ) и относительный содержание воды ( RWC ) при полной насыщенности (после 48 часов под вакуумом) в образцах цельной, ранней и поздней древесины Показаны средние значения (±SE) ( n =5).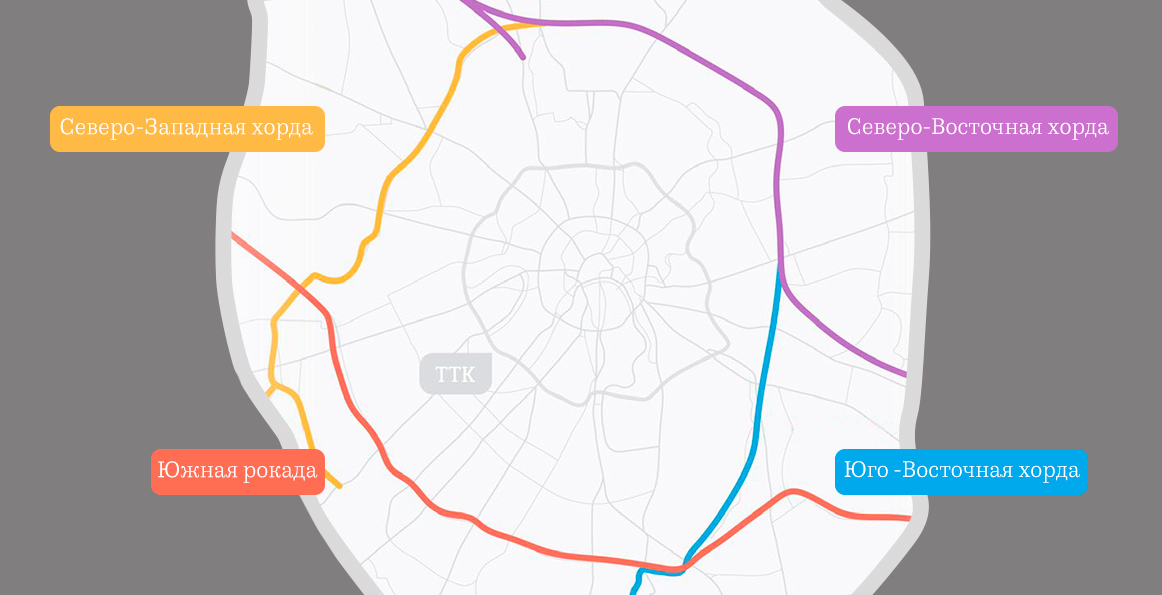 Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05.
Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05.
| Плотность (G CM -3 ) | MC (%) | V Cellwall (%) | V H 2 O ( %) | В Газ (%) | RWC (%) | |||||
| Цельная древесина | 0,39±0.02 A | 182 ± 12 A | 25,3 ± 1,2 А | 69,3 ± 2.1 A | 69.3 ± 2.1 A | 5,4 ± 1,4 A | 93 ± 2 A | |||
| Ранчание | 0,26 ± 0,02 B | 316 ± 18 B | 16,7 ± 1,0 В | 79,9 ± 1,0 млрд. | 3,4 ± 1,3 A | 96 ± 2 A | 96-2 A | 96 ± 2 A | ||
| Противодействие | 0,60 ± 0,01 С | 92 ± 3 C | 39,6 ± 0,7 C | 55,4 ± 1.1 C | 5.1 ± 1,0 A | 92 ± 2 A |
| Плотность (G CM -3 ) | MC (%) | V Cellwall ( %) | V H 2 O (%) | GAS (%) | (%) | RWC (%) | ||
| цельноугольный | 0. 39 ± 0,02 A | 182 ± 12 A | 182 ± 12,3 ± 1,2 A | 69,3 ± 2.1 A | 5,4 ± 1,4 A | 93 ± 2 A | ||
| Ранчание | 0,26 ± 0,02 B | 316 ± 18 B | 16,7 ± 1,0 B | 79,9 ± 1,0 B | 99,9 ± 1,4 ± 1,3 A96 ± 2 A | 96 ± 2 A | ||
| 0,60 ± 0,01 С | 92 ± 3 C | 39,6 ± 0,7 C | 55,4±1,1 в | 5,1±1,0 а | 92±2 а |
Плотность древесины (сухая масса / сырой объем), содержание влаги ( MC ), объем, занимаемый материалом клеточной стенки ( V клеточная стенка ), объем, занимаемый водой1 ( 9086 H 2 O ), объем, занимаемый газом ( V газ ), и относительное содержание воды ( RWC) в условиях полного вакуума через 4 часа при насыщении 90 образцы цельной, ранней и поздней древесины Показаны средние значения (±SE) ( n =5). Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05.
Значения в столбце с одной и той же буквой существенно не отличаются: 90 185 P 90 186 = 0,05.
| Плотность (G CM -3 ) | MC (%) | V Cellwall (%) | V H 2 O ( %) | В Газ (%) | RWC (%) | |||||
| Цельная древесина | 0,39±0.02 A | 182 ± 12 A | 25,3 ± 1,2 А | 69,3 ± 2.1 A | 69.3 ± 2.1 A | 5,4 ± 1,4 A | 93 ± 2 A | |||
| Ранчание | 0,26 ± 0,02 B | 316 ± 18 B | 16,7 ± 1,0 В | 79,9 ± 1,0 млрд. | 3,4 ± 1,3 A | 96 ± 2 A | 96-2 A | 96 ± 2 A | ||
| Противодействие | 0,60 ± 0,01 С | 92 ± 3 C | 39,6 ± 0,7 C | 55,4 ± 1.1 C | 5.1 ± 1,0 A | 92 ± 2 A |
| Плотность (G CM -3 ) | MC (%) | V Cellwall ( %) | V H 2 O (%) | GAS (%) | (%) | RWC (%) | ||
| цельноугольный | 0. 39 ± 0,02 A 39 ± 0,02 A | 182 ± 12 A | 182 ± 12,3 ± 1,2 A | 69,3 ± 2.1 A | 5,4 ± 1,4 A | 93 ± 2 A | ||
| Ранчание | 0,26 ± 0,02 B | 316 ± 18 B | 16,7 ± 1,0 B | 79,9 ± 1,0 B | 99,9 ± 1,4 ± 1,3 A96 ± 2 A | 96 ± 2 A | ||
| 0,60 ± 0,01 С | 92 ± 3 C | 39,6 ± 0,7 C | 55,4±1,1 в | 5,1±1,0 а | 92±2 а |
Измеренное относительное содержание влаги в ранней и поздней древесине ( RWC ) и расчетная потеря электропроводности в процентах ( PLC ) по результатам летних измерений на поле Значение поля зимы Ψ ствола составило –0,5±0,1 МПа. Минимальные значения поля летнего Ψ ствола в предрассветный и полуденный период составили –1,0±0,1 МПа и –1,4±0,1 МПа соответственно. Соответствующий летний Ψ лист в полдень был равен –1. 7±0,1 МПа ( n =6).
7±0,1 МПа ( n =6).
| Измеренный RWC зима | Измеренный RWC лето | Оценочных PLC от измеренного RWC | Сметных PLC от измеренного ф ствола | |
| Earlywood | 79 ± 2 | 75 ± 4 | 22 ± 8 | 16 ± 3 | 91 092
| Latewood | 65 ± 2 | 57 ± 1 | 56 ± 3 | 42 ± 2 | 91 092
| Измеренный RWC зима | Измеренный RWC лето | Оценочное PLC от измеренного RWC | Оценочное PLC от измеряется Ψ ствол | |
| Earlywood | 79 ± 2 | 75 ±4 | 22±8 | 16±3 |
| Поздняя древесина | 65±2 | 57±1 | 56±3 | 42±2 |
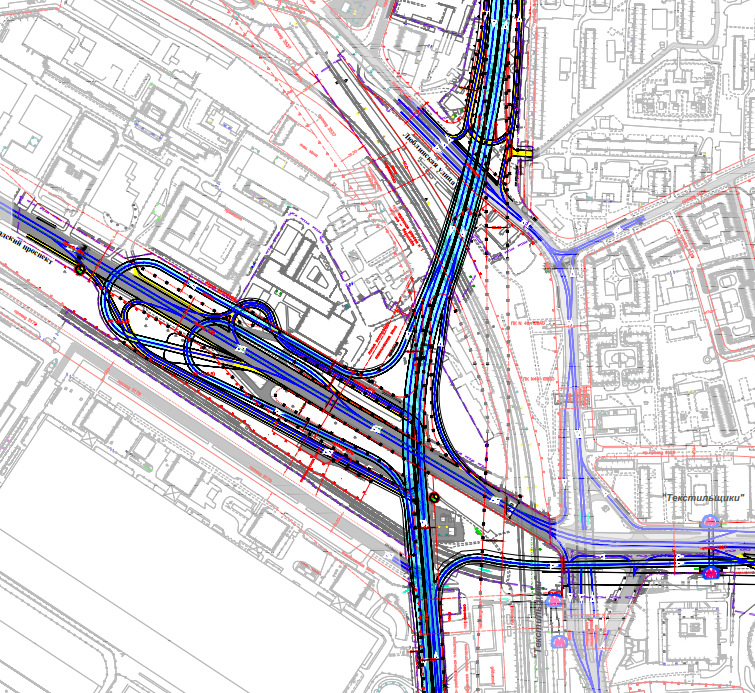
Измеренное относительное содержание влаги в ранней и поздней древесине ( RWC ) и расчетная потеря электропроводности в процентах ( PLC ) по результатам летних измерений на поле Значение поля зимы Ψ ствола составило –0,5±0,1 МПа. Минимальные значения поля летнего Ψ ствола в предрассветный и полуденный период составили –1,0±0,1 МПа и –1,4±0,1 МПа соответственно. Соответствующий летний Ψ лист в полдень был равен –1.7±0,1 МПа ( n =6).
| Измеренный RWC зима | Измеренный RWC лето | Оценочных PLC от измеренного RWC | Сметных PLC от измеренного ф ствола | |
| Earlywood | 79 ± 2 | 75 ± 4 | 22 ± 8 | 16 ± 3 | 91 092
| Latewood | 65 ± 2 | 57 ± 1 | 56 ± 3 | 42 ± 2 | 91 092
| Измеренный RWC зима | Измеренный RWC лето | Оценочное PLC от измеренного RWC | Оценочное PLC от измеряется Ψ ствол | |
| Earlywood | 79 ± 2 | 75 ±4 | 22±8 | 16±3 |
| Поздняя древесина | 65±2 | 57±1 | 56±3 | 42±2 |
Ссылки
Ольха NN , Sperry JS, Pockman WT.
1996
. Эмболия корней и стебля ксилемы, устьичная проводимость и тургор листьев в популяциях Acer grandidentatum вдоль градиента влажности почвы.Экология
105
,293
–301.Алони Р .
2001
. Фолиарные и осевые аспекты сосудистой дифференцировки: гипотезы и доказательства.Журнал правил выращивания растений
20
,22
–34.Баннан МВт .
1967
. Антиклинальные деления и длина клеток в камбии хвойных.Журнал лесных товаров
17
,63
–69.Барахас-Моралес J .
1985
. Структурные различия древесины между деревьями двух тропических лесов Мексики.Журнал IAWA
6
,355
–364.
Библия EJ .
1969
. Переходные вариации и отношения между свойствами в годичных кольцах сосны Лоблолли.Древесина Наука и технология
3
,14
–24.Бодиг J , Джейн Б.А.
1982
. Механика древесины и древесных композитов . Нью-Йорк, штат Нью-Йорк Компания Van Nostrand Reinhold.Болтон AJ , Петти JA.
1978
.Модель, описывающая осевое течение жидкости через хвойную древесину.Древесина Наука и технология
12
,37
–48.Карлквист S .
1985
. Васицентрическая трахеида как механизм выживания в засухе.Алисо
11
,37
–68. Cochard H , Peiffer M, Le Gall K, Granier A.
1997
. Контроль развития гидравлического сопротивления ксилемы и уязвимости к эмболии у Fraximus excelsior L.влияет на водные отношения.Журнал экспериментальной ботаники
48
,655
–663.Каун DJ .
1975
. Вариации размеров трахеид в стволе 26-летней лучистой сосны.Аппита
28
,237
–245.Кромби DS , Хипкинс М.Ф., Милберн Дж.А.
1985
. Проникновение газа в ямочные мембраны ксилемы Rhododendron как причина акустически обнаруживаемой соковой кавитации.Австралийский журнал физиологии растений
12
,445
–453.Де Корт I .
1993
. Взаимосвязь между количеством заболони, процентным содержанием поздней древесины, содержанием влаги и жизнеспособностью кроны пихты Дугласа.
Журнал IAWA
14
,413
–427.Domec J-C , Gartner BL.
2001
. Кавитация и водоаккумулирующая способность в сегментах ксилемы ствола взрослых и молодых деревьев пихты Дугласа.Деревья
15
,204
–214.Domec J-C , Gartner BL.
2002
и . Связанные с возрастом и положением изменения гидравлической и механической дисфункции ксилемы: вывод критериев проектирования структуры древесины дугласовой пихты.Физиология деревьев
22
,91
–104.Domec J-C , Gartner BL.
2002
б .Влияние изменения структуры ямки, окаймленной пихтой Дугласа, и ее функционирования на гидравлику дерева. Материалы Национального собрания СВС 2001 года. Денвер, Ко,351
–356.
Эдвардс WRN , Джарвис П.Г.
1982
. Взаимосвязь между содержанием воды, водным потенциалом и водопроницаемостью в стволах хвойных.Растения, клетки и окружающая среда
5
,271
–277.Эллмор GS , Эверс FW.
1986
. Поток жидкости в самом внешнем приросте ксилемы кольцевидного дерева, Ulmus americana .Американский журнал ботаники
73
,1771
–1774.Эверс FW , Карлтон М.Р., Фишер Дж.Б., Тайри МТ.
1997
. Диаметры сосудов в корнях по сравнению со стеблями тропических лиан и других форм роста.Журнал IAWA
18
,261
–279.Фенгель Д .
1972
. Структура и функции мембраны в окаймленных хвойными породами ямах.
Holzforschung
26
,1
–9.Gibson AC , HW Calkin, Nobel PS.
1985
. Гидравлическая проводимость и структура ксилемы у трахеидоносных растений.Бюллетень IAWA
6
,293
–302.Гилмор AR , Бойс С.Г., Райкер Р.А.
1966
. Связь удельного веса сосны лоблолли с факторами окружающей среды в южном Иллинойсе.Лесоведение
12
,399
–405.Гольдштейн Г , Андраде Д.Л., Мейнцер Ф.С., Холбрук Н.М., Кавелье Дж., Джексон П., Селис А.
1998
. Запасы стволовой воды и суточные закономерности использования воды деревьями в пологе тропических лесов.Растения, клетки и окружающая среда
21
,397
–406. Грегори SG , Петти Дж. А.
А.
1973
. Клапанное действие окаймленных ямок у хвойных.Журнал экспериментальной ботаники
24
,763
–767.Hacke UG , Sperry JS, Pockman WT, Davis SD, McCulloh KA.
2001
. Тенденции изменения плотности и структуры древесины связаны с предотвращением имплозии ксилемы за счет отрицательного давления.Экология
126
,457
–461.Харгрейв KR , Колб К.Дж., Эверс Ф.В., Дэвис С.Д.
1994
. Диаметр канала и вызванная засухой эмболия у Salvia mellifera Green (Labiatae).Новый фитолог
126
,695
–705.Холбрук Нью-Мексико .
1995
. Хранение стволовой воды. В: Gartner BL, изд. Стебли растений: физиология и функциональная морфология .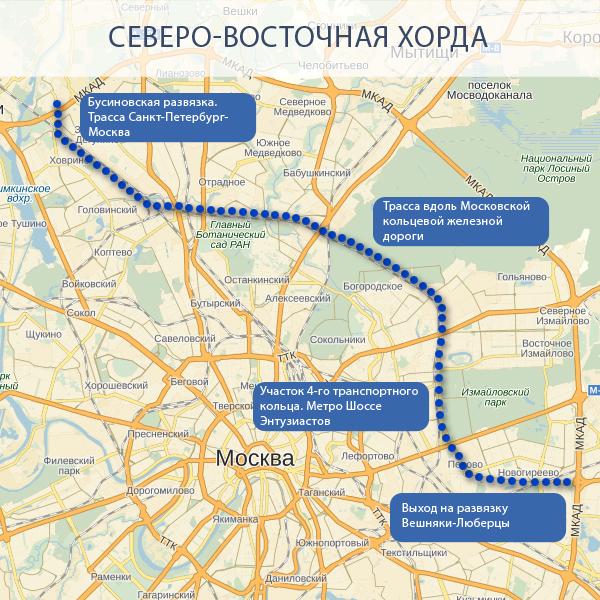 Сан-Диего: Academic Press,
Сан-Диего: Academic Press,105
–124.Келлог RM , Sastry CBR, Wellwood RW.
1975
. Связь между составом клеточной стенки и плотностью клеточной стенки.Древесное волокно
7
,170
–177.Кеннеди РВ .
1961
. Изменчивость и периодичность летней древесины у некоторых дугласовых пихт второго роста.Tappi Journal
44
,161
–165.Кеннеди RW , Уоррен В.Г.
1969
. Изменение физических и химических свойств пихты Дугласа внутри дерева. В: Вторая всемирная консультация по селекции лесных деревьев , Vol. 1 . Вашингтон, округ Колумбия: Международный союз организаций по исследованию леса,393
–417. Ларсон PR .
1962
. Биологический подход к качеству древесины.Tappi Journal
45
,443
–448.Ларсон PR .
1994
. Сосудистый камбий: развитие и строение . Нью-Йорк: Springer-Verlag.Лебуржуа Ф .
2000
Климатические сигналы в ранней древесине и общей ширине годичных колец корсиканской сосны из западной Франции.Летопись лесоведения
57
,155
–164.Ло Гулло MA , Саллео С., Пьячери ЕС, Россо Р.
1995
.Взаимосвязь между уязвимостью к ксилемной эмболии и размерами каналов ксилемы у молодых деревьев Quercus cerris .Растения, клетки и окружающая среда
18
,661
–670. Лутц Дж .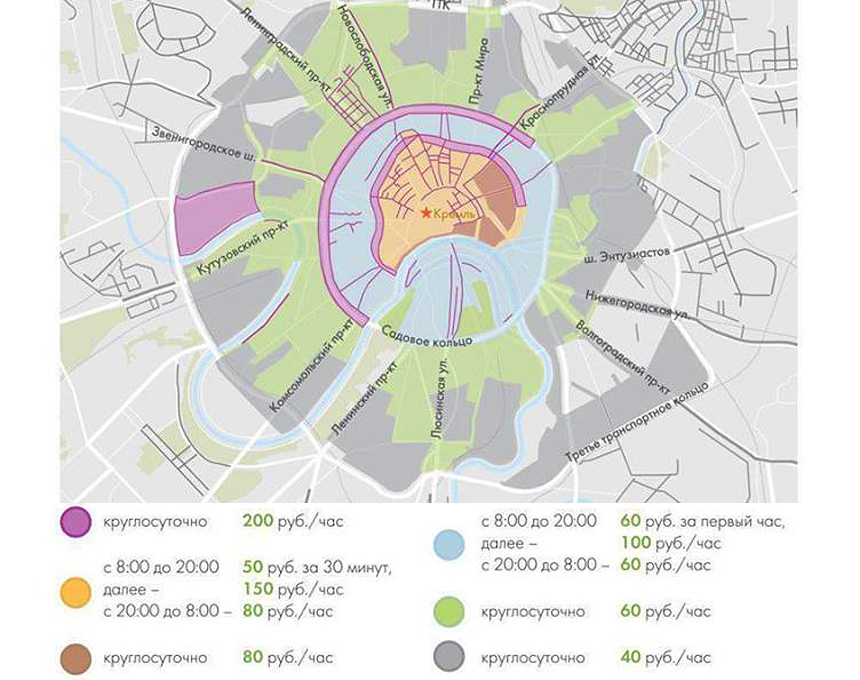
1964
. Влияние скорости роста на свойства шпона хвойных пород.Журнал лесных товаров
24
,94
–102.Мегроу РА .
1985
. Показатели качества древесины сосны лоболли . Атланта, Джорджия: TAPPI Press.Matzner SL , Райс К.Дж., Ричардс Дж.Х.
2001
. Внутривидовые вариации кавитации ксилемы живого дуба внутри ( Quercus wislizenii A. DC.).Журнал экспериментальной ботаники
52
,783
–789.Макмиллан CW .
1968
. Морфологические характеристики древесины сосны лоблоловой в зависимости от удельного веса, скорости роста и расстояния от сердцевины.Древесина Наука и технология
2
,166
–176. Мотт Л , Лес Г. , Шалер С.
, Шалер С.
2002
. Механические свойства отдельных волокон сосны южной. Часть II. Сравнение волокон ранней и поздней древесины по высоте и молодости.Древесина и волокно
34
,212
–220.Паншин AJ , де Зеев К.
1980
. Учебник по технологии древесины , 4-е изд. Нью-Йорк: Макгроу-Хилл.Парк S .
1984
. Структура «противоположного дерева». II. Вариабельность диаметра и толщины стенки трахеиды, ширины кольца и процента поздней древесины.Мокузай Гаккаиси
30
,110
–116.Петти ТА .
1972
. Аспирация окаймленных ям в хвойной древесине.Труды Королевского общества, серия B
181
,395
-406. Петти JA , Puritch GS.
1970
. эффекты сушки на структуру и проницаемость древесины Abies Grandis .Вуд Наука и техника
4
,140
-154.Покман WT , Sperry JS.
2000
. Уязвимость ксилемной кавитации и распространение растительности пустыни Сонора.Американский журнал ботаники
87
,1287
–1299.Польж Н .
1973
. Facteurs ecologiques et qualité du bois.Летопись лесоведения
30
,307
–328.Расбанд Вт .
1996
. Изображение Национального института здравоохранения. Версия 1.60 . Национальные институты здоровья, Бетесда, Мэриленд.Рихтер Н .
1976
. Водный статус растения — экспериментальные данные. В: Ланге О.Л., Каппен, Шульце Э.Д., ред. Вода и растения . Берлин: Springer,
В: Ланге О.Л., Каппен, Шульце Э.Д., ред. Вода и растения . Берлин: Springer,42
–58.Саллео S , Хинкли Т.М., Кикута С.Б., Ло Гулло М.А., Вейлгони П., Юнанд Т.М., Рихтер Х.
1992
. Метод индуцирования ксилемных эмболов in situ : эксперименты с полевым деревом.Растения, клетки и окружающая среда
15
,491
–497.SAS Institute Inc.
1997
. Программное обеспечение SAS версии 6.12. SAS Institute Inc.: Кэри, Северная Каролина.Schulte PJ , Гибсон AC.
1988
. Гидравлическая проводимость и анатомия трахеид у шести видов современных семенных растений.Canadian Journal of Botany
66
,1073
–1079.Сиау JF .
1984
. Транспортные процессы в древесине .Берлин: Springer-Verlag.
Транспортные процессы в древесине .Берлин: Springer-Verlag.Sperry JS , Saliendra NZ.
1994
. Внутри- и межрастительные вариации кавитации ксилемы у Betula occidentalis .Растения, клетки и окружающая среда
17
,1233
–1241.Сперри JS , Тайри МТ.
1988
. Механизм ксилемной эмболии, вызванной водным стрессом.Физиология растений
88
,581
–587.Сперри JS , Тайри МТ.
1990
. Ксилемная эмболия, вызванная водным стрессом, у трех видов хвойных.Растения, клетки и окружающая среда
13
,427
–436.Spicer R , Gartner BL.
1998
и . Каким образом голосеменная ветвь принимает гидравлический статус главного стебля, когда берет верх над лидером?Растения, клетки и окружающая среда
21
,1063
–1070.
Spicer R , Gartner BL.
1998
б . Гидравлические свойства ветвей и половинок пихты Дугласа ( Pseudotsuga menziesii ) по отношению к сжатой древесине.Физиология деревьев
18
,777
–784.Stratton L , Goldstein G, Meinzer FC.
2000
. Вместимость стволовых вод и эффективность водного транспорта: их функциональное значение в сухом гавайском лесу.Растения, клетки и окружающая среда
23
,99
–106Takjima T .
1967
. Рост деревьев и свойства древесины . Токио: Сельскохозяйственный факультет Токийского университета.Timell TE .
1986
. Сжатая древесина голосеменных растений , Vol. 1. Берлин: Springer-Verlag.
1. Берлин: Springer-Verlag.Тайри MT , Дэвис С.Т., Кочард Х.
1994
. Биофизические перспективы эволюции ксилемы: есть ли компромисс между гидравлической эффективностью и уязвимостью к дисфункциям?IAWA Journal
15
,335
–360Tyree MT , Ewers FW.
1991
. Обзор Тэнсли № 34. Гидравлическая архитектура деревьев и других древесных растений.Новый фитолог
119
,345
–360.Тайри МТ , Sperry JS.
1988
. Работают ли древесные растения вблизи точки катастрофической дисфункции ксилемы, вызванной динамическим водным стрессом: ответ модели.Физиология растений
88
,574
–580.Тайри МТ , Ян С.
1990
. Влагоемкость стеблей Thuja, Tsuga и Acer , измеренная по изотермам обезвоживания.
Влагоемкость стеблей Thuja, Tsuga и Acer , измеренная по изотермам обезвоживания.Планта
182
,420
–426.Waring RH , Работающий SW.
1998
. Анализ экосистем в множественном масштабе . Сан-Диего Пресс.Вестинг АХ .
1968
. Формирование и функция компрессионной древесины у голосеменных растений. II.Ботаническое обозрение
34
,51
–78.Ёсидзава N , Киёмия М., Идей Т.
1987
.Вариации длины трахеид и морфологические изменения кончиков трахеид, связанные с развитием компрессионной древесины.Древесина Наука и технология
21
,1
–10.Циммерманн МХ .
1983
.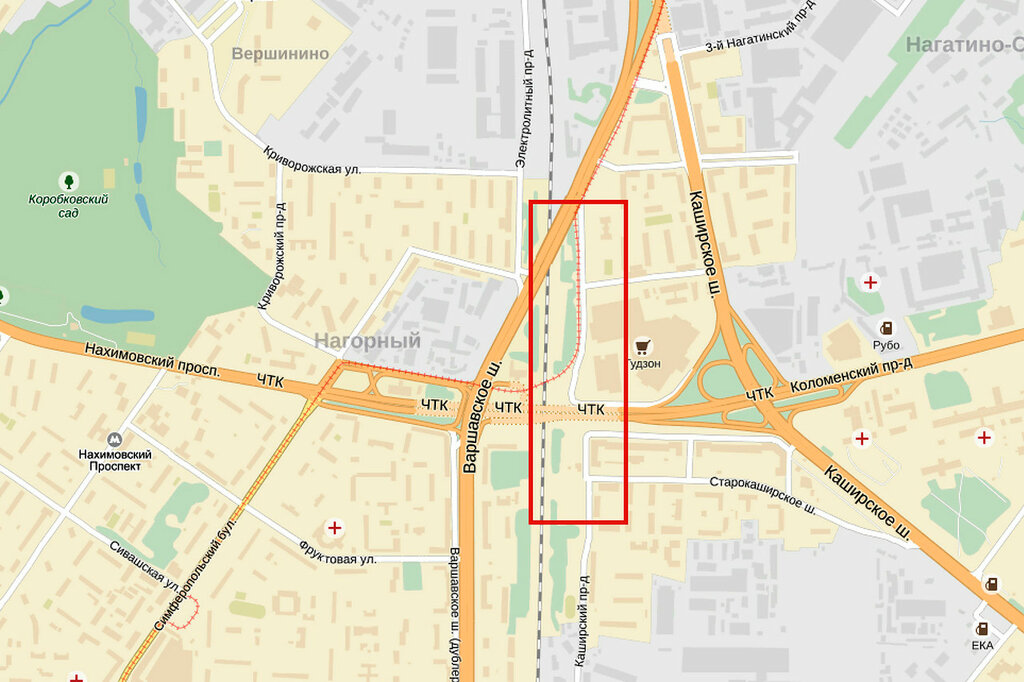 Структура ксилемы и восхождение сока . Берлин: Springer-Verlag.
Структура ксилемы и восхождение сока . Берлин: Springer-Verlag.Зобель BH , Ван Буйтенен JP .
1989
. Различия между деревьями и внутри них.В: Изменчивость древесины: ее причины и борьба с ней . Нью-Йорк: Springer-Verlag.%PDF-1.2 % 2620 0 объект > эндообъект внешняя ссылка 2620 165 0000000016 00000 н 0000003656 00000 н 0000003841 00000 н 0000003874 00000 н 0000003933 00000 н 0000005384 00000 н 0000005563 00000 н 0000005633 00000 н 0000005752 00000 н 0000005845 00000 н 0000006027 00000 н 0000006206 00000 н 0000006378 00000 н 0000006481 00000 н 0000006672 00000 н 0000006863 00000 н 0000006972 00000 н 0000007084 00000 н 0000007269 00000 н 0000007383 00000 н 0000007487 00000 н 0000007607 00000 н 0000007717 00000 н 0000007830 00000 н 0000007928 00000 н 0000008043 00000 н 0000008152 00000 н 0000008281 00000 н 0000008399 00000 н 0000008506 00000 н 0000008627 00000 н 0000008735 00000 н 0000008899 00000 н 0000009069 00000 н 0000009202 00000 н 0000009325 00000 н 0000009505 00000 н 0000009672 00000 н 0000009807 00000 н 0000009925 00000 н 0000010101 00000 н 0000010202 00000 н 0000010365 00000 н 0000010531 00000 н 0000010635 00000 н 0000010716 00000 н 0000010845 00000 н 0000010949 00000 н 0000011045 00000 н 0000011219 00000 н 0000011355 00000 н 0000011467 00000 н 0000011566 00000 н 0000011682 00000 н 0000011801 00000 н 0000011911 00000 н 0000012037 00000 н 0000012191 00000 н 0000012304 00000 н 0000012446 00000 н 0000012643 00000 н 0000012747 00000 н 0000012855 00000 н 0000013041 00000 н 0000013143 00000 н 0000013332 00000 н 0000013459 00000 н 0000013585 00000 н 0000013749 00000 н 0000013859 00000 н 0000013968 00000 н 0000014105 00000 н 0000014280 00000 н 0000014419 00000 н 0000014505 00000 н 0000014610 00000 н 0000014726 00000 н 0000014817 00000 н 0000014919 00000 н 0000015018 00000 н 0000015115 00000 н 0000015212 00000 н 0000015310 00000 н 0000015408 00000 н 0000015506 00000 н 0000015604 00000 н 0000015703 00000 н 0000015804 00000 н 0000015903 00000 н 0000016002 00000 н 0000016101 00000 н 0000016200 00000 н 0000016299 00000 н 0000016398 00000 н 0000016497 00000 н 0000016596 00000 н 0000016695 00000 н 0000016794 00000 н 0000016893 00000 н 0000016992 00000 н 0000017091 00000 н 0000017190 00000 н 0000017289 00000 н 0000017388 00000 н 0000017489 00000 н 0000017588 00000 н 0000017687 00000 н 0000017786 00000 н 0000017885 00000 н 0000017984 00000 н 0000018083 00000 н 0000018182 00000 н 0000018281 00000 н 0000018380 00000 н 0000018479 00000 н 0000018578 00000 н 0000018677 00000 н 0000018776 00000 н 0000018875 00000 н 0000018974 00000 н 0000019073 00000 н 0000019172 00000 н 0000019271 00000 н 0000019370 00000 н 0000019469 00000 н 0000019568 00000 н 0000019667 00000 н 0000019768 00000 н 0000019867 00000 н 0000019966 00000 н 0000020065 00000 н 0000020164 00000 н 0000020263 00000 н 0000020362 00000 н 0000020461 00000 н 0000020560 00000 н 0000020659 00000 н 0000020758 00000 н 0000020857 00000 н 0000020956 00000 н 0000021055 00000 н 0000021154 00000 н 0000021253 00000 н 0000021352 00000 н 0000021453 00000 н 0000021552 00000 н 0000021652 00000 н 0000021751 00000 н 0000021850 00000 н 0000021949 00000 н 0000022048 00000 н 0000022276 00000 н 0000022458 00000 н 0000022513 00000 н 0000022712 00000 н 0000023496 00000 н 0000024432 00000 н 0000024641 00000 н 0000025433 00000 н 0000025652 00000 н 0000026451 00000 н 0000027310 00000 н 0000027390 00000 н 0000003976 00000 н 0000005360 00000 н трейлер ] >> startxref 0 %%EOF 2621 0 объект > эндообъект 2622 0 объект [ 2623 0 Р ] эндообъект 2623 0 объект > /Ф 2697 0 Р >> эндообъект 2624 0 объект > эндообъект 2783 0 объект > ручей HV[lU4aSJ4 LjTj,qNMwٖv-Fbdh5Qj$FD
Лос-Аламосская национальная лаборатория: Домашняя страница MCNP
Лос-Аламосская национальная лаборатория
РЕФЕРАТ
MCNP — универсальный M onte C arlo N — P артикул, код которого
может использоваться для переноса нейтронов, фотонов, электронов или связанных нейтронов/фотонов/электронов. Конкретные области применения включают, помимо прочего, радиационную защиту.
и дозиметрия, радиационная защита, радиография, медицинская физика, безопасность ядерной критичности,
Проектирование и анализ детектора, каротаж ядерных нефтяных скважин, проектирование мишени ускорителя,
Проектирование ядерных и термоядерных реакторов, дезактивация и вывод из эксплуатации.
Код обрабатывает произвольную трехмерную конфигурацию материалов.
в геометрических ячейках, ограниченных поверхностями первой и второй степени и
эллиптические торы четвертой степени.
Конкретные области применения включают, помимо прочего, радиационную защиту.
и дозиметрия, радиационная защита, радиография, медицинская физика, безопасность ядерной критичности,
Проектирование и анализ детектора, каротаж ядерных нефтяных скважин, проектирование мишени ускорителя,
Проектирование ядерных и термоядерных реакторов, дезактивация и вывод из эксплуатации.
Код обрабатывает произвольную трехмерную конфигурацию материалов.
в геометрических ячейках, ограниченных поверхностями первой и второй степени и
эллиптические торы четвертой степени.
Обычно используются данные точечного поперечного сечения, хотя также доступны групповые данные.
Для нейтронов все реакции, указанные в конкретной оценке поперечного сечения (например, ENDF/B-VI).
учитываются. Тепловые нейтроны описываются как моделями свободного газа, так и S(альфа,бета) моделями.
Для фотонов код учитывает некогерентное и когерентное рассеяние, возможность
флуоресцентное излучение после фотоэлектрического поглощения, поглощение в парном образовании с локальным
испускание аннигиляционного излучения и тормозное излучение. Модель с непрерывным замедлением
используется для переноса электронов, включая позитроны, k-лучи и тормозное излучение, но не
включать внешние или самоиндуцированные поля.
Модель с непрерывным замедлением
используется для переноса электронов, включая позитроны, k-лучи и тормозное излучение, но не
включать внешние или самоиндуцированные поля.
Важные стандартные функции, которые делают MCNP очень универсальным и простым в использовании, включают мощный общий источник, источник критичности и поверхностный источник; как геометрические, так и итоговые плоттеры; а богатая коллекция методов уменьшения дисперсии; гибкая структура подсчета; и обширный сбор данных поперечного сечения.
MCNP содержит множество гибких счетчиков: поверхностный ток и поток, объемный поток (длина дорожки), точечные или кольцевые детекторы, нагрев частиц, нагрев делением, подсчет амплитуды импульса для энергии или осаждение заряда, подсчеты сеток и подсчеты радиографии.
Основные моменты MCNP
См. вкладку НОВОСТИ MCNP для получения информации о:
Спецвыпуск журнала Nuclear Technology
Деятельность по поддержке сайта MCNP в LANL
Симпозиум пользователей MCNP
Дополнения к справочной коллекции MCNP (13 сентября 2020 г.
 )
)План и требования SQA MCNP6, LA-UR-20-26666
Дополнения к справочной коллекции MCNP (07.11.2019)
Около 40 отчетов и документов по безопасности по критичности, неструктурированная сетка, постдокторские и студенческие работы NCS, статистика.
Дополнения к справочной коллекции MCNP (05.02.2018)
Примечания к выпуску MCNP6.2
Дополнения к справочной коллекции MCNP (21 ноября 2017 г.)
Руководство пользователя MCNP6.2
Сборка MCNP6.2
Тестирование версии MCNP6.2
Примечание о ядерных данных для MCNP6 (28 сентября 2017 г.)
Как указано в ссылках ниже, предыдущие данные, опубликованные с
MCNP6.1 и MCNP6.1.1 были следующие проблемы:
(1) Отсутствуют (n,g) данные о производстве водорода в файлах ACE, 1001,80c — 1001,86c
(2) Ошибки в данных теплового рассеяния SiO2 S(a,b), sio2,30t — sio2. 36t
36t
(3) Ошибки в данных теплового рассеяния h-zr.27t S(a,b) при 1200K
Эти вопросы данных обсуждаются в отчетах на веб-сайте MCNP, в
Справочная коллекция, раздел «Информация о выпуске версии MCNP6.2»:
* Выпуск MCNP6.2 и Whisper-1.1
— Руководство для пользователей NCS, LA-UR-17-24260
* Что нового в MCNP6.2 и Whisper-1.1, LA-UR-17-27992
* Проверка MCNP6.2 для приложений безопасности ядерной критичности,
LA-UR-17-24406
* Проверка MCNP6.2 для
Приложения безопасности ядерной критичности, LA-UR-17-23822.
* Список доступных таблиц данных ACE, LA-UR-17-20709.
* Изменения данных для выпуска MCNP6.2, LA-UR-17-21486, LA-UR-17-20703
MCNP6.Релиз 2 включает в себя исправленные файлы данных. Они обсуждаются в ссылках выше и в примечаниях к выпуску MCNP6.2.
MCNP® и Monte Carlo N-Particle® являются зарегистрированными товарными знаками, принадлежащими
Triad National Security, LLC, менеджер и оператор Лос-Аламоса
Национальная лаборатория. Любое использование третьим лицом таких зарегистрированных знаков должно
быть должным образом отнесены к Triad National Security, LLC, включая
использование обозначения ® в зависимости от ситуации.Любые вопросы по лицензированию,
надлежащее использование и/или надлежащее указание торговых марок Triad National Security, LLC
следует направлять по адресу [email protected].
Любое использование третьим лицом таких зарегистрированных знаков должно
быть должным образом отнесены к Triad National Security, LLC, включая
использование обозначения ® в зависимости от ситуации.Любые вопросы по лицензированию,
надлежащее использование и/или надлежащее указание торговых марок Triad National Security, LLC
следует направлять по адресу [email protected].
Cytochrome C — Chemistry LibreTexts
Всем клеткам требуется энергия в виде АТФ, аденозинтрифосфата, для запуска основных метаболических процессов, необходимых для выживания. Когда аэробный организм переваривает пищу, глюкоза (C 6 H 12 O 6 ) в процессе гликолиза расщепляется на две молекулы пирувата.Восстановленные переносчики электронов, NADH и FADH 2 образуются как побочный продукт этой реакции.
NAD + получает два электрона (2e — ) и ион водорода (H + ), образуя NADH. Точно так же FAD принимает два иона водорода (2H + ) и два электрона (2e —) с образованием FADH 2 . Эти электроны из NADH и FADH 2 затем входят в комплексы электрон-транспортной цепи. Цепь переноса электронов (ЭТЦ) представляет собой серию белков, переносящих электроны, расположенных во внутренней мембране митохондрий — электростанции клетки (рис. 1).В митохондриальную мембрану встроен ряд окислительно-восстановительных белков, которые действуют как провод, перемещая электроны через ЭТЦ. Через серию окислительно-восстановительных реакций эти белки перекачивают протоны (H + ) из митохондриального матрикса в межмембранное пространство. Накопление протонов в межмембранном пространстве создает электрохимический градиент, который обеспечивает синтез АТФ ферментом АТФ-синтазой. Цепь переноса электронов производит в общей сложности 34 молекулы АТФ, которые клетка может использовать для осуществления своих метаболических процессов для выживания. 1
Эти электроны из NADH и FADH 2 затем входят в комплексы электрон-транспортной цепи. Цепь переноса электронов (ЭТЦ) представляет собой серию белков, переносящих электроны, расположенных во внутренней мембране митохондрий — электростанции клетки (рис. 1).В митохондриальную мембрану встроен ряд окислительно-восстановительных белков, которые действуют как провод, перемещая электроны через ЭТЦ. Через серию окислительно-восстановительных реакций эти белки перекачивают протоны (H + ) из митохондриального матрикса в межмембранное пространство. Накопление протонов в межмембранном пространстве создает электрохимический градиент, который обеспечивает синтез АТФ ферментом АТФ-синтазой. Цепь переноса электронов производит в общей сложности 34 молекулы АТФ, которые клетка может использовать для осуществления своих метаболических процессов для выживания. 1
Рисунок 1 . Цепь переноса электронов
Это общий обзор электрон-транспортной цепи (ETC). ЭТЦ представляет собой серию из четырех белковых комплексов: НАДН-дегидрогеназы, сукцинатдегидрогеназы, цитохрома bc 1 и цитохрома с оксидазы, встроенных во внутреннюю мембрану митохондрий. По мере переноса электронов через эти белковые комплексы в межмембранном пространстве митохондрий накапливается протонный (Н + ) градиент.АТФ-синтаза использует этот протонный градиент, установленный ETC, для синтеза энергии в форме АТФ.
ЭТЦ представляет собой серию из четырех белковых комплексов: НАДН-дегидрогеназы, сукцинатдегидрогеназы, цитохрома bc 1 и цитохрома с оксидазы, встроенных во внутреннюю мембрану митохондрий. По мере переноса электронов через эти белковые комплексы в межмембранном пространстве митохондрий накапливается протонный (Н + ) градиент.АТФ-синтаза использует этот протонный градиент, установленный ETC, для синтеза энергии в форме АТФ.
Рис. 2. Полноразмерная ленточная структура цитохрома C (PDB 3cyt). 3 Это биологическая сборка одного мономера цитохрома с.Внутри центрального гема находится ион железа, показанный оранжевым цветом. Гем ковалентно связан с белком через дисульфидные связи, что стало возможным благодаря повторяющемуся мотиву Cys-XY-Cys-His, обнаруженному в цитохромах. | Цитохром с является важным компонентом электрон-транспортной цепи для синтеза АТФ (рис. 2). Цитохром с является водорастворимым электрон-транспортным белком, свободно связанным с внутренней мембраной митохондрий.В электрон-транспортной цепи цитохром с переносит по одному электрону за раз через свою гемовую группу от третьего комплекса электрон-транспортной цепи, цитохрома bc 1 , к четвертому комплексу электрон-транспортной цепи, цитохром с-оксидазе. 2 Цитохром с содержит металлический центр гемового железа, который необходим для его функции. Во время процесса переноса электронов это гемовое железо взаимопревращается между состояниями окисления Fe 3+ и Fe 2+ , что позволяет принимать и отдавать электроны. 4 Когда цитохром с находится в окисленной форме, электрон переносится от комплекса цитохрома bc 1 к гему Fe 3+ , восстанавливая его до Fe 2+ . |

 Наконец, цитохром с отдает электрон конечному переносчику электронов ЭТЦ, цитохром с оксидазе. В этот момент центр железа вернется к своей степени окисления Fe 3+ .
Наконец, цитохром с отдает электрон конечному переносчику электронов ЭТЦ, цитохром с оксидазе. В этот момент центр железа вернется к своей степени окисления Fe 3+ . Железометаллический центр цитохрома с представляет собой октаэдрическую геометрию из-за координации шести лигандов вокруг центрального иона железа (рис. 3).Октаэдрическая геометрия предпочтительна для цитохрома с, потому что каждый из 6 лигандов, богатых электронами, способствует стабилизации положительно заряженного иона металлического железа. Железо гема, показанное оранжевым цветом, координируется четырьмя атомами азота жесткого квадратного плоского порфиринового кольца и двумя аксиальными лигандами: атомом серы остатка метионина и атомом азота гистидинимидазольного кольца. Порфириновое кольцо цитохрома с считается тетрадентатным хелатирующим лигандом, поскольку четыре атома азота порфиринового кольца связываются с центральным железом, образуя стабильный металлоорганический комплекс. Основываясь на хелатном эффекте, этот сайт связывания тетрадентатного лиганда является более энтропийно благоприятным по сравнению с сродством монодентатного лиганда к тому же иону металла. 5
Основываясь на хелатном эффекте, этот сайт связывания тетрадентатного лиганда является более энтропийно благоприятным по сравнению с сродством монодентатного лиганда к тому же иону металла. 5
Лиганды цитохрома с подходят на основании теории жестких мягких кислотных оснований. (HSAB). HSAB классифицирует кислоты и основания как твердые, мягкие или пограничные. Мягкие кислоты и основания больше и легко поляризуются, в то время как твердые кислоты и основания меньше и менее поляризуются. Виды, которые находятся между жесткими и мягкими, считаются пограничными. 6 Исходя из HSAB, логично, что пограничное железо координируется с азотом гистидинимидазольного кольца (пограничное основание), атомами азота порфиринового кольца (пограничное основание) и серой метионина (пограничное основание). мягкая основа).
Рисунок 3 . Железо-металлический центр цитохрома С 3
На рис. 3А показана вся структура цитохрома с, а на рис. 3В показана увеличенная врезка металлического центра железа гема, необходимого для функции переноса электронов цитохрома с.Хим железо, показанное в апельсине, координируется четырьмя атомами азота жесткого квадрата планарного кольца порфирина (синий), а также два осевых лиганды: атом серы метионина (желтый) и атом азота гистидина имидазола (синий ).
3В показана увеличенная врезка металлического центра железа гема, необходимого для функции переноса электронов цитохрома с.Хим железо, показанное в апельсине, координируется четырьмя атомами азота жесткого квадрата планарного кольца порфирина (синий), а также два осевых лиганды: атом серы метионина (желтый) и атом азота гистидина имидазола (синий ).
Центр металлического железа цитохрома с представляет собой октаэдрическую геометрию из-за координации шести лигандов вокруг центрального иона железа (рис. 3). Октаэдрическая геометрия предпочтительна для цитохрома с, потому что каждый из 6 лигандов, богатых электронами, способствует стабилизации положительно заряженного иона металлического железа.Железо гема, показанное оранжевым цветом, координируется четырьмя атомами азота жесткого квадратного плоского порфиринового кольца и двумя аксиальными лигандами: атомом серы остатка метионина и атомом азота гистидинимидазольного кольца. Порфириновое кольцо цитохрома с считается тетрадентатным хелатирующим лигандом, поскольку четыре атома азота порфиринового кольца связываются с центральным железом, образуя стабильный металлоорганический комплекс. Основываясь на хелатном эффекте, этот сайт связывания тетрадентатного лиганда является более энтропийно благоприятным по сравнению с сродством монодентатного лиганда к тому же иону металла. 5
Основываясь на хелатном эффекте, этот сайт связывания тетрадентатного лиганда является более энтропийно благоприятным по сравнению с сродством монодентатного лиганда к тому же иону металла. 5
Лиганды цитохрома с подходят на основании теории жестких мягких кислотных оснований. (ХСАБ). HSAB классифицирует кислоты и основания как твердые, мягкие или пограничные. Мягкие кислоты и основания больше по размеру и легко поляризуются, а жесткие кислоты и основания меньше по размеру и менее поляризуемы. Виды, которые находятся между жесткими и мягкими, считаются пограничными. 6 Исходя из HSAB, логично, что пограничное железо координируется с азотом гистидинимидазольного кольца (пограничное основание), атомами азота порфиринового кольца (пограничное основание) и серой метионина (пограничное основание). мягкая основа).
Хотя металлический центр железа гема изменяет степень окисления во время процесса переноса электронов, цитохром с всегда принимает октаэдрическую низкоспиновую геометрию независимо от степени окисления железа. 7 Эта геометрия является предпочтительной на основе энергии стабилизации поля лиганда (LFSE). LFSE представляет собой полную энергию d-электронов металлического комплекса относительно теоретического барицентра. Формула для определения LFSE показана в уравнении 1 ниже, где x = количество d-электронов на низкоэнергетических орбиталях t 2g , а y = количество d-электронов на высокоэнергетических e g орбиталях.
7 Эта геометрия является предпочтительной на основе энергии стабилизации поля лиганда (LFSE). LFSE представляет собой полную энергию d-электронов металлического комплекса относительно теоретического барицентра. Формула для определения LFSE показана в уравнении 1 ниже, где x = количество d-электронов на низкоэнергетических орбиталях t 2g , а y = количество d-электронов на высокоэнергетических e g орбиталях.
Рис. 4. Диаграмма LFSE разделения низкоспинового Fe 3+ и Fe 2+
Диаграммы расщепления электронов и расчеты для низкоспиновых Fe 3+ и Fe 2+ показаны на рисунках A и B соответственно. Более отрицательные значения LFSE для низкоспинового железа по сравнению с высокоспиновым железом указывают на то, что эта конформация с низким спином является более энергетически выгодной и, следовательно, представляет собой конфигурацию, которую примет комплекс.
Рисунок 5 . Диаграммы расщепления LFSE высокоспиновых Fe 3+ и Fe 2+
Диаграммы расщепления LFSE высокоспиновых Fe 3+ и Fe 2+
Теоретические диаграммы электронного расщепления и расчеты для высокоспиновых Fe 3+ и Fe 2+ показаны на рисунках A и B соответственно. Однако значения LFSE более отрицательны для низкоспинового железа, чем для высокоспинового железа, независимо от степени окисления. Следовательно, центр металлического железа гема цитохрома с всегда будет иметь низкоспиновую октаэдрическую геометрию независимо от степени окисления железа.
В состоянии с низким спином d-электроны спариваются на орбиталях с низкой энергией t 2g , прежде чем занять орбитали с более высокой энергией e g , что повышает стабильность. В высокоспиновом состоянии d-электроны по отдельности занимают обе орбитали t 2g и e g независимо от энергии орбиталей, а затем спариваются. Чем отрицательнее LFSE, тем стабильнее комплекс. Значения LFSE для низкоспинового железа более отрицательны, чем значения LFSE для высокоспинового железа, независимо от степени окисления железа. Таким образом, центр металлического железа цитохрома с всегда будет принимать более энергетически выгодную низкоспиновую конформацию.
Таким образом, центр металлического железа цитохрома с всегда будет принимать более энергетически выгодную низкоспиновую конформацию.
В процессе переноса электронов гемовое железо цитохрома с циклически переключается между состояниями окисления +2 и +3. Поскольку цитохром с принимает электрон от третьего комплекса электрон-транспортной цепи, цитохрома bc 1, , железометаллический центр Fe 3+ восстанавливается до Fe 2+ . Когда цитохром с отдает этот электрон конечному комплексу электрон-транспортной цепи, цитохром с-оксидазе, степень окисления Fe 3+ восстанавливается.Однако, несмотря на это изменение степени окисления железа, координация гема не меняется; ион железа «заперт на месте» и претерпевает минимальную реорганизацию.
Восстановительный потенциал также способствует электрон-транспортной функции цитохрома с в ЭТЦ. Восстановительный потенциал (E o ’) — это тенденция химического вещества приобретать электроны и, следовательно, восстанавливаться. Чем положительнее восстановительный потенциал, тем больше склонность этого химического вещества принимать электрон и восстанавливаться.Восстановительный потенциал цитохрома bc 1 (комплекс III ЭТЦ) составляет 0,194 В. Цитохром bc 1 отдает один электрон окисленной форме цитохрома с (Fe 3+ ), восстанавливая железо цитохрома с на одна степень окисления, чтобы стать Fe 2+ . Восстановительный потенциал для цитохрома с равен 0,254 В. Восстановительный потенциал для цитохром с оксидазы (комплекс IV ЦЭТ) составляет 0,562 В. цитохром с в его окисленную форму (Fe 3+ ).Комплексы электрон-транспортной цепи расположены в порядке возрастания окислительно-восстановительного потенциала (каждый комплекс имеет более высокое сродство к электронам, чем предыдущий), что направляет поток электронов к последнему комплексу электрон-транспортной цепи, цитохром-С-оксидазе.
Чем положительнее восстановительный потенциал, тем больше склонность этого химического вещества принимать электрон и восстанавливаться.Восстановительный потенциал цитохрома bc 1 (комплекс III ЭТЦ) составляет 0,194 В. Цитохром bc 1 отдает один электрон окисленной форме цитохрома с (Fe 3+ ), восстанавливая железо цитохрома с на одна степень окисления, чтобы стать Fe 2+ . Восстановительный потенциал для цитохрома с равен 0,254 В. Восстановительный потенциал для цитохром с оксидазы (комплекс IV ЦЭТ) составляет 0,562 В. цитохром с в его окисленную форму (Fe 3+ ).Комплексы электрон-транспортной цепи расположены в порядке возрастания окислительно-восстановительного потенциала (каждый комплекс имеет более высокое сродство к электронам, чем предыдущий), что направляет поток электронов к последнему комплексу электрон-транспортной цепи, цитохром-С-оксидазе.
E o ’донор < E o ’ белок переноса электрона/центр < E o ’ акцептор: Должен быть истинным, чтобы реакция была спонтанной!
В заключение, цитохром с является важным белком переноса электронов, который перемещает электроны между комплексами III и IV ETC.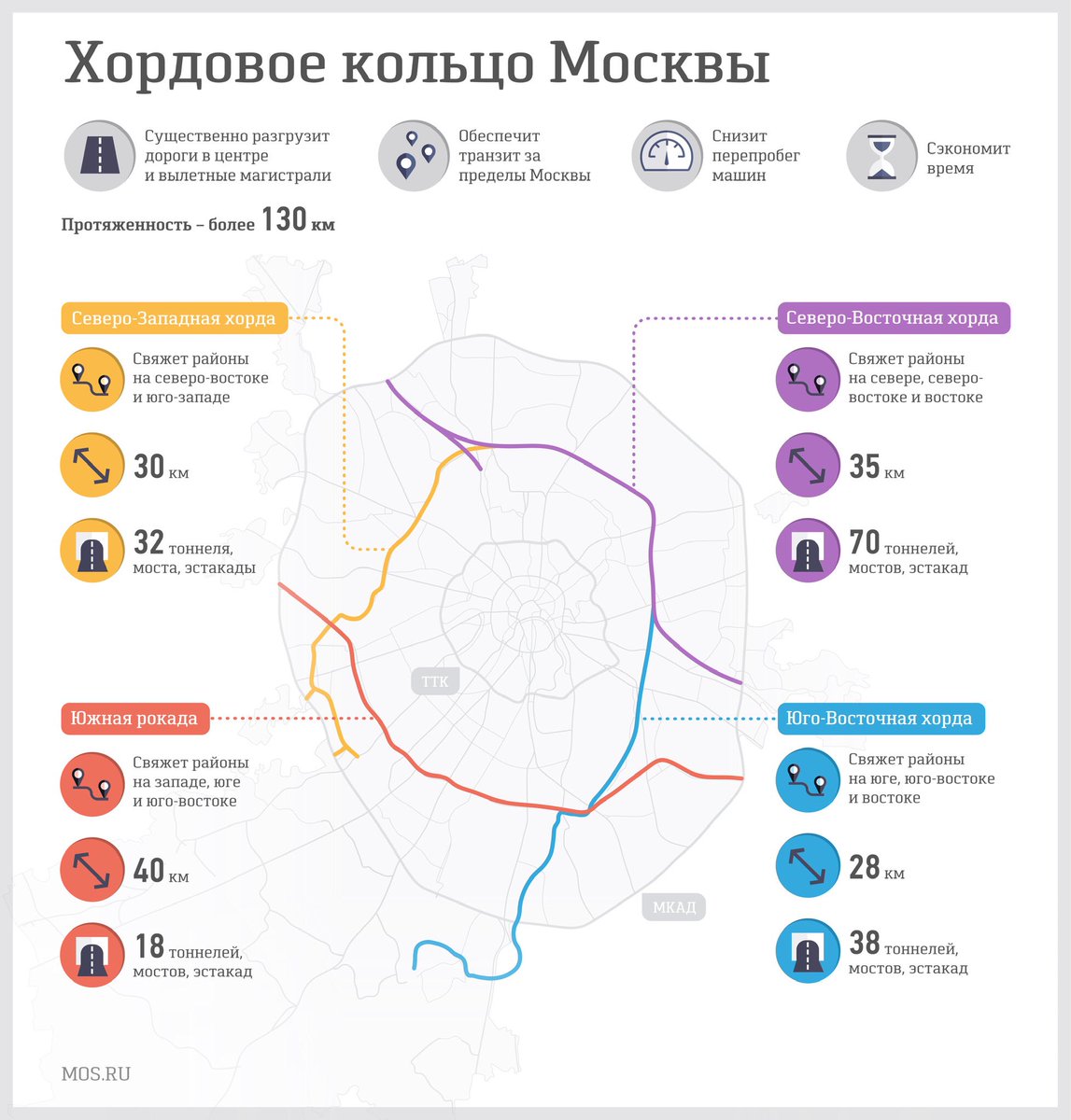 Металлический центр железа гема легко взаимопревращается между Fe 3+ и Fe 2+ , что позволяет принимать и отдавать электроны. Если бы этого переноса электронов не происходило, АТФ, необходимая для питания многих метаболических процессов, таких как движение мышц, синтез ДНК и активный транспорт, не производилась бы. Следовательно, цитохром с является важным компонентом, способствующим выработке этого АТФ посредством переноса его электронов на цитохром С-оксидазу, которая обеспечивает клетки энергией, необходимой им для выполнения этих жизненно важных процессов, необходимых для выживания. 9
Металлический центр железа гема легко взаимопревращается между Fe 3+ и Fe 2+ , что позволяет принимать и отдавать электроны. Если бы этого переноса электронов не происходило, АТФ, необходимая для питания многих метаболических процессов, таких как движение мышц, синтез ДНК и активный транспорт, не производилась бы. Следовательно, цитохром с является важным компонентом, способствующим выработке этого АТФ посредством переноса его электронов на цитохром С-оксидазу, которая обеспечивает клетки энергией, необходимой им для выполнения этих жизненно важных процессов, необходимых для выживания. 9
Источники
- Alberts, B.; Джонсон, А .; Льюис, Дж, et al. Молекулярная биология клетки . 4-й ред.; Нью-Йорк. Наука гирлянды; 2002. Цепи переноса электронов и их белковые насосы. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK26904/ .
- Лефинтер, утра.; Нельсон, Д.Л.; Кокс, М.М. лейнингских принципов биохимии ; Высшее образование Macmillan: Houndmills, 2017.
 .
. - Такано, т.; Дикерсон, Р.Э. Редокс-конформационные изменения в очищенном цитохроме с тунца. проц. Натл. акад. науч. США. 1980. 77, 6371-6375.
- Роут-Мэлоун, Р.М. Бионеорганическая химия – краткий курс; Wiley & Sons: Хобокен, Нью-Джерси, 2007 г.
- Синьшань, К. и Кэри, Дж. (1999). Роль гема в структурной организации цитохрома с, изученная методом полусинтеза. Биохимия, 38 (48), 15944-15951.
- Химия LibreTexts: Жесткие и мягкие кислоты и основания
- Сильва, Дж.младший; Уильямс, Р.Дж.П. Биологическая химия элементов: неорганическая химия жизни; Издательство Оксфордского университета: Оксфорд, 2001.
- Бертини, И. Биологическая неорганическая химия: структура и реакционная способность ; Университетские научные книги: Саусалито, Калифорния, 2007. .
- Эрециньска, М.; Сильвер, И.А. АТФ и функция мозга. Журнал мозгового кровотока и метаболизма.